

In silico APC/C substrate discovery reveals cell cycle-dependent degradation of UHRF1 and other chromatin regulators
- PMID: 33306668
- PMCID: PMC7758050
- DOI: 10.1371/journal.pbio.3000975
In silico APC/C substrate discovery reveals cell cycle-dependent degradation of UHRF1 and other chromatin regulators
Abstract
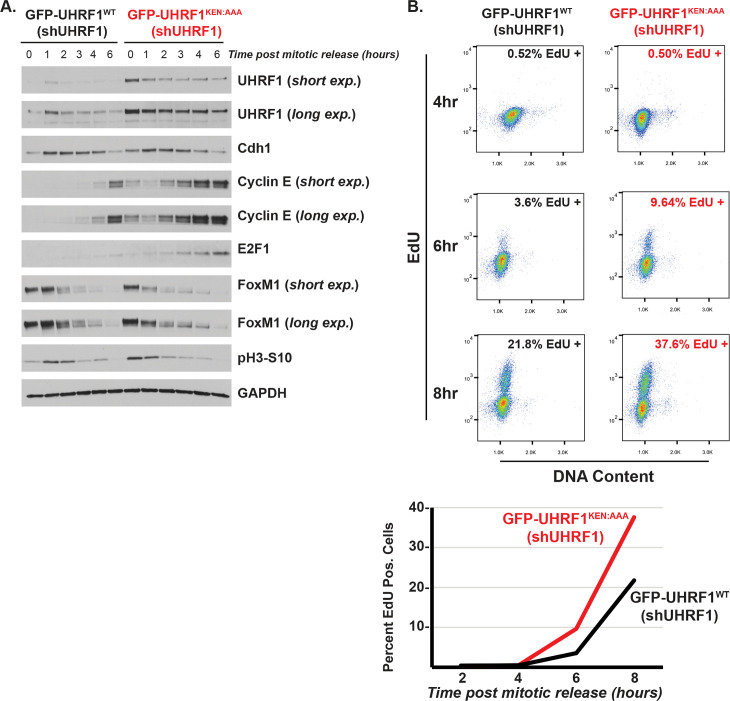
The anaphase-promoting complex/cyclosome (APC/C) is an E3 ubiquitin ligase and critical regulator of cell cycle progression. Despite its vital role, it has remained challenging to globally map APC/C substrates. By combining orthogonal features of known substrates, we predicted APC/C substrates in silico. This analysis identified many known substrates and suggested numerous candidates. Unexpectedly, chromatin regulatory proteins are enriched among putative substrates, and we show experimentally that several chromatin proteins bind APC/C, oscillate during the cell cycle, and are degraded following APC/C activation, consistent with being direct APC/C substrates. Additional analysis revealed detailed mechanisms of ubiquitylation for UHRF1, a key chromatin regulator involved in histone ubiquitylation and DNA methylation maintenance. Disrupting UHRF1 degradation at mitotic exit accelerates G1-phase cell cycle progression and perturbs global DNA methylation patterning in the genome. We conclude that APC/C coordinates crosstalk between cell cycle and chromatin regulatory proteins. This has potential consequences in normal cell physiology, where the chromatin environment changes depending on proliferative state, as well as in disease.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: NGB is a consultant for Cullgen, Inc.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
