

Structural basis of cytoplasmic NaV1.5 and NaV1.4 regulation
- PMID: 33306788
- PMCID: PMC7953540
- DOI: 10.1085/jgp.202012722
Structural basis of cytoplasmic NaV1.5 and NaV1.4 regulation
Abstract
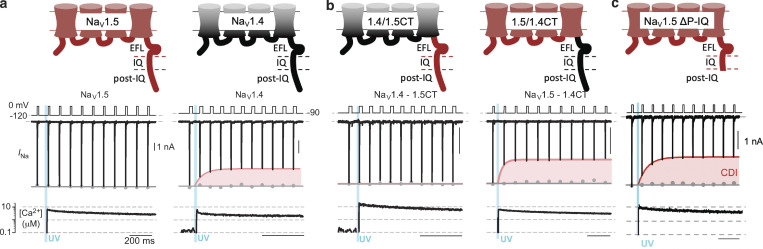
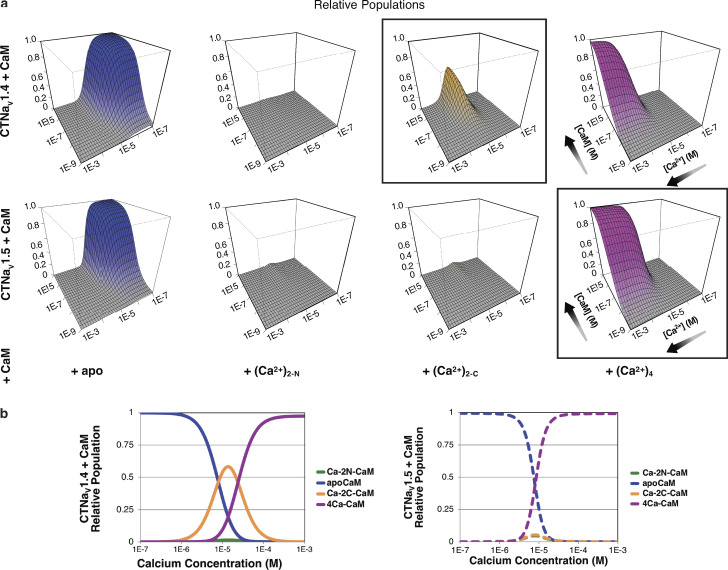
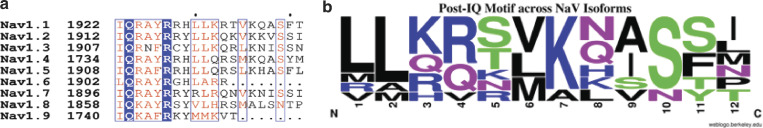
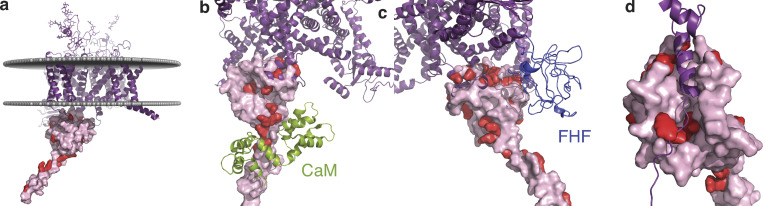
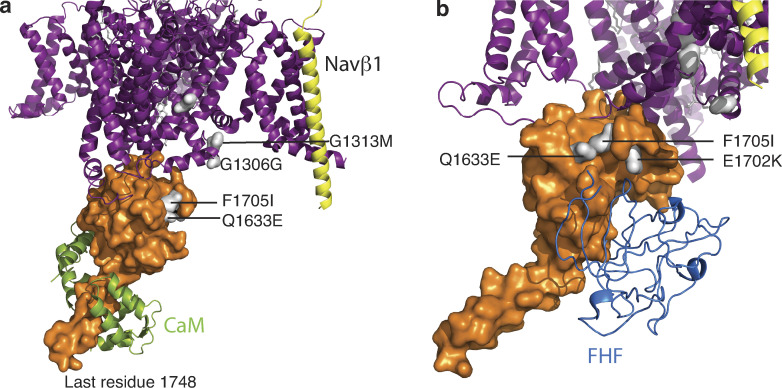
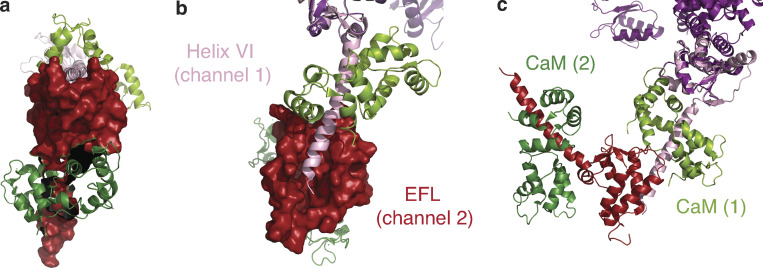
Voltage-gated sodium channels (NaVs) are membrane proteins responsible for the rapid upstroke of the action potential in excitable cells. There are nine human voltage-sensitive NaV1 isoforms that, in addition to their sequence differences, differ in tissue distribution and specific function. This review focuses on isoforms NaV1.4 and NaV1.5, which are primarily expressed in skeletal and cardiac muscle cells, respectively. The determination of the structures of several eukaryotic NaVs by single-particle cryo-electron microscopy (cryo-EM) has brought new perspective to the study of the channels. Alignment of the cryo-EM structure of the transmembrane channel pore with x-ray crystallographic structures of the cytoplasmic domains illustrates the complementary nature of the techniques and highlights the intricate cellular mechanisms that modulate these channels. Here, we review structural insights into the cytoplasmic C-terminal regulation of NaV1.4 and NaV1.5 with special attention to Ca2+ sensing by calmodulin, implications for disease, and putative channel dimerization.
© 2020 Nathan et al.
Figures
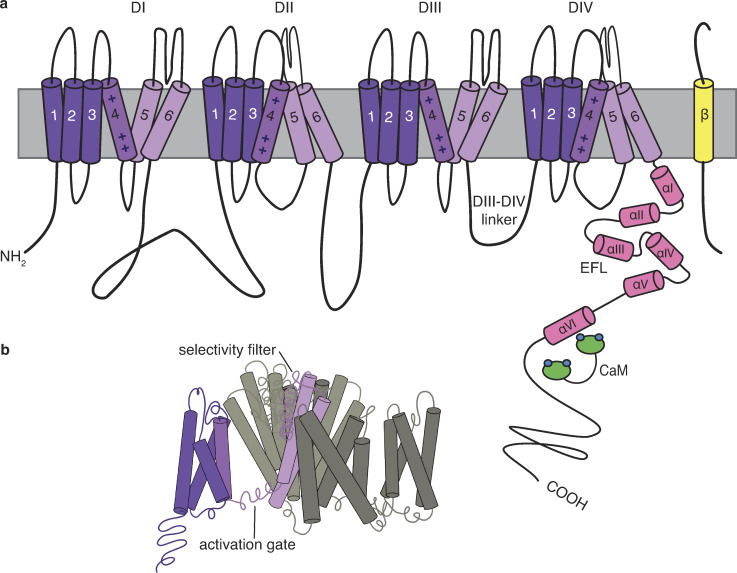
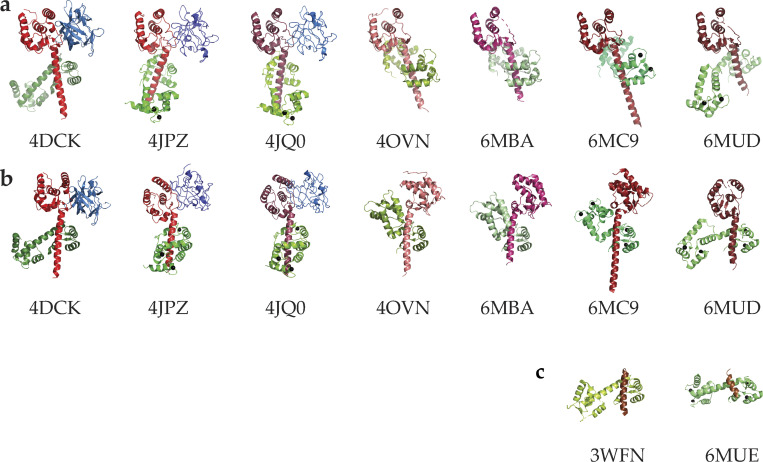
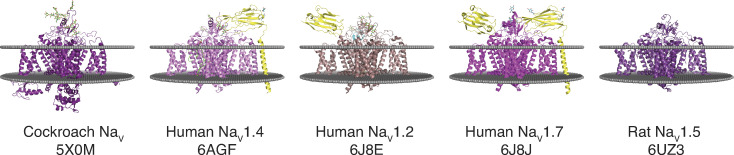
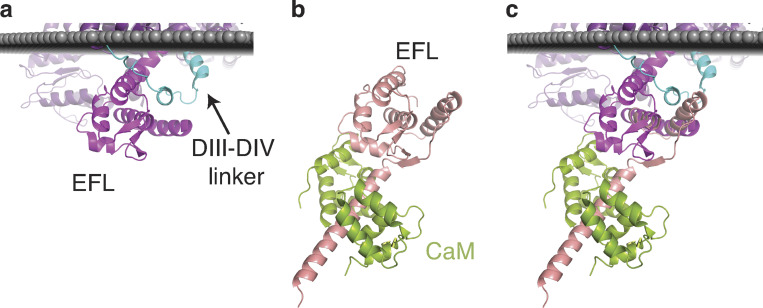
References
-
- Agullo-Pascual, E., Lin X., Leo-Macias A., Zhang M., Liang F.X., Li Z., Pfenniger A., Lübkemeier I., Keegan S., Fenyö D., et al. 2014. Super-resolution imaging reveals that loss of the C-terminus of connexin43 limits microtubule plus-end capture and NaV1.5 localization at the intercalated disc. Cardiovasc. Res. 104:371–381. 10.1093/cvr/cvu195 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
