

Close to the edge: Heterochromatin at the nucleolar and nuclear peripheries
- PMID: 33307247
- PMCID: PMC7855492
- DOI: 10.1016/j.bbagrm.2020.194666
Close to the edge: Heterochromatin at the nucleolar and nuclear peripheries
Abstract
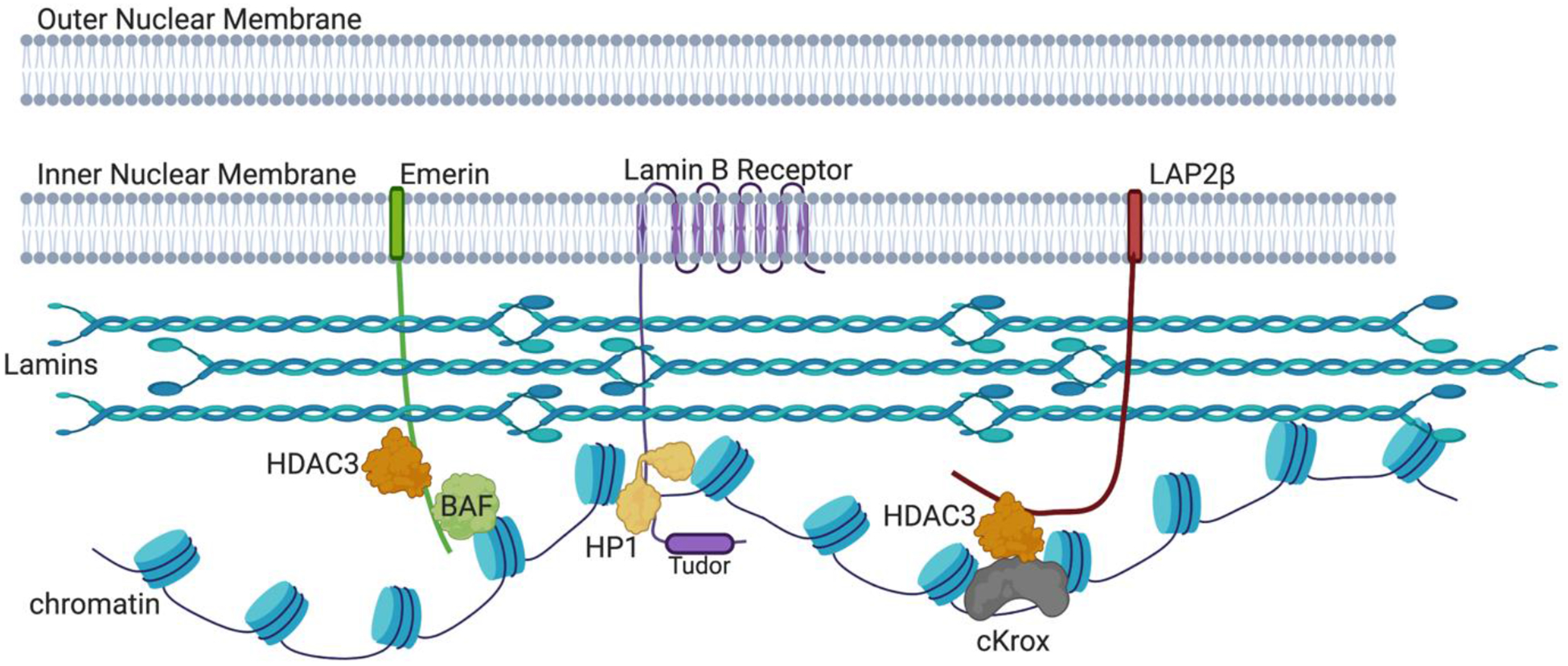
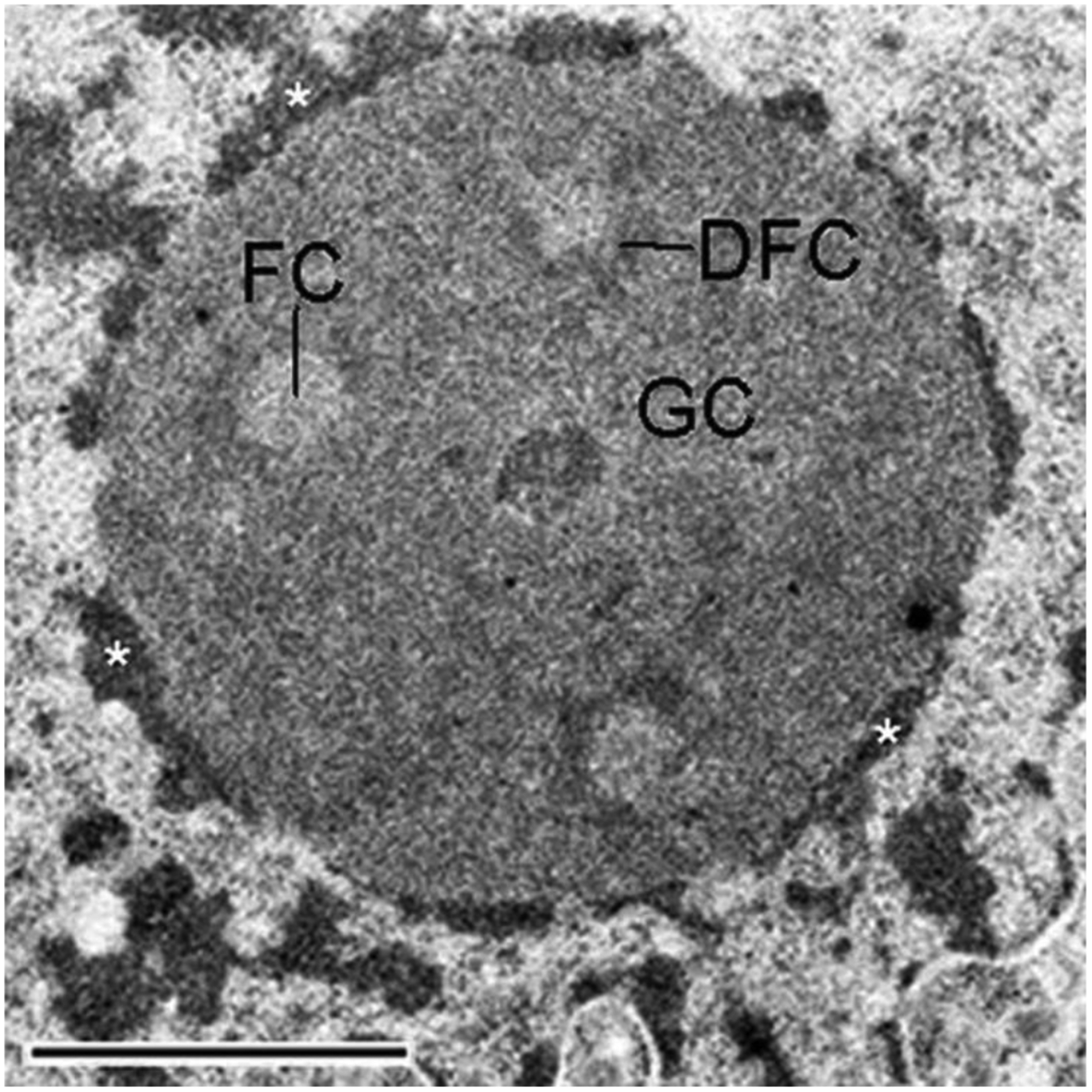
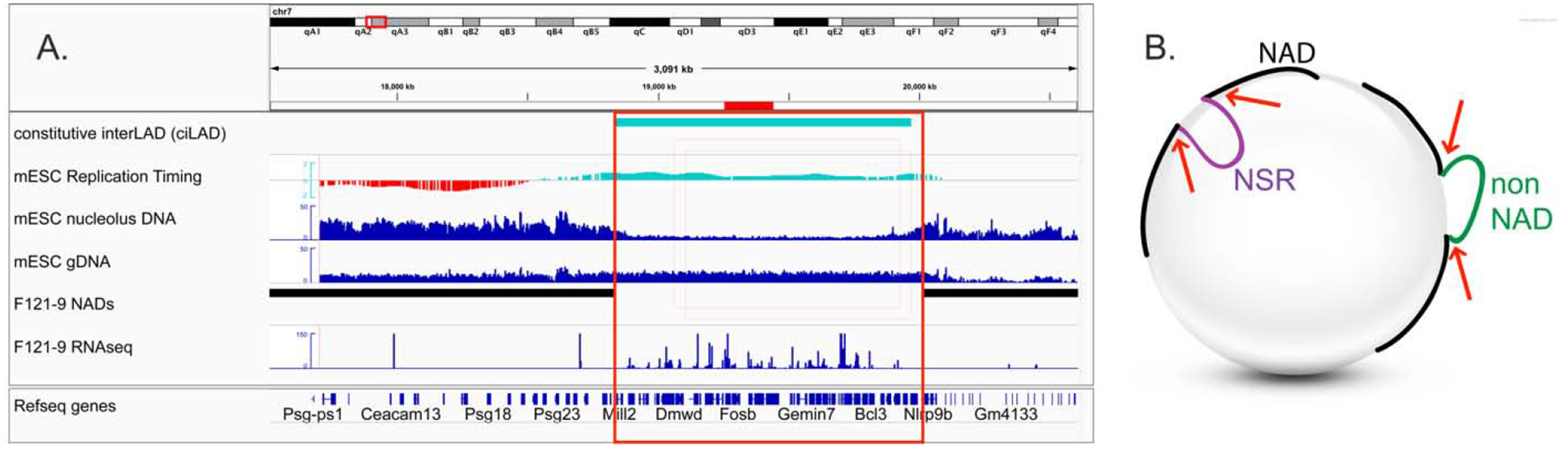
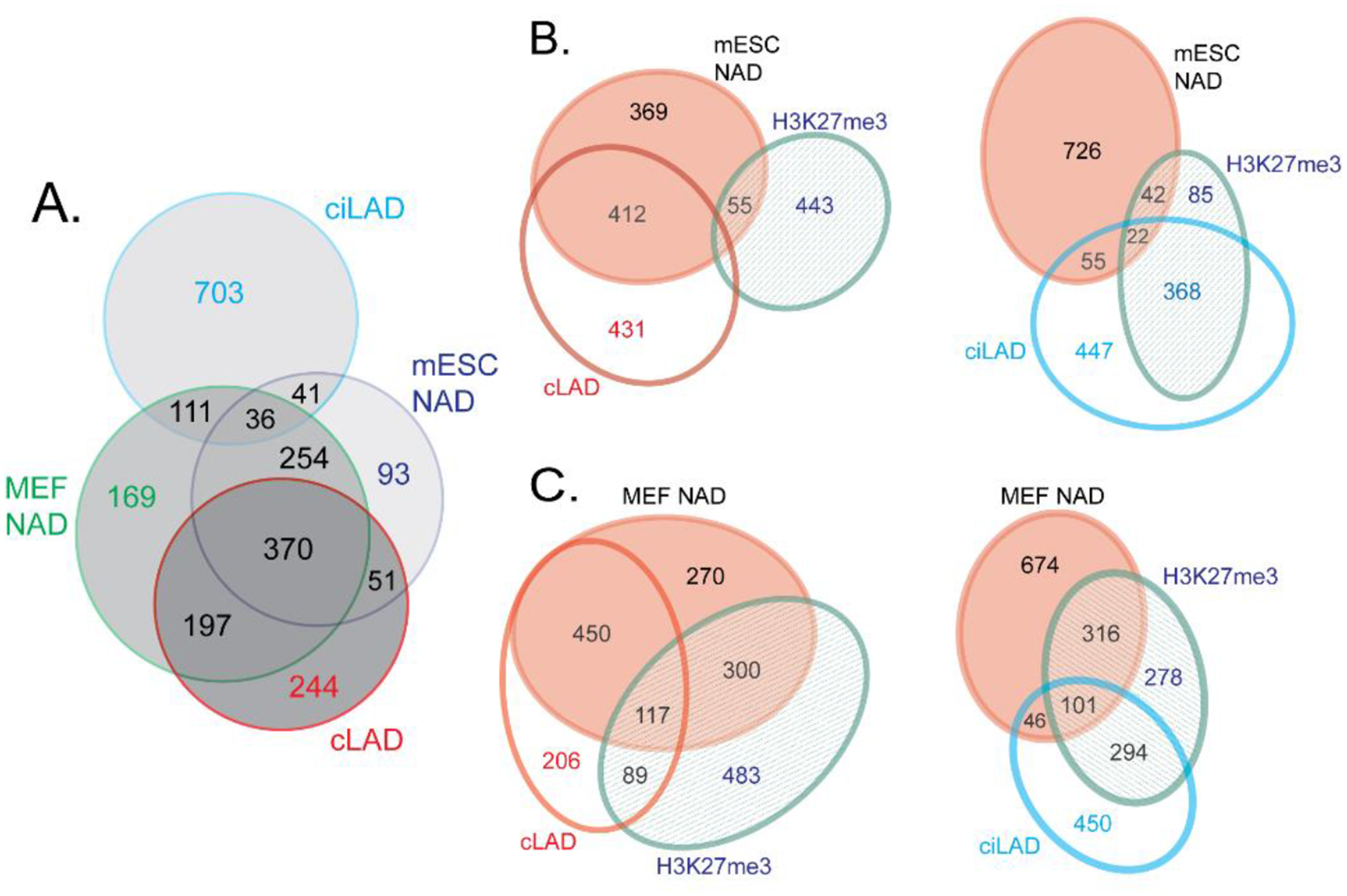
Chromatin is a dynamic structure composed of DNA, RNA, and proteins, regulating storage and expression of the genetic material in the nucleus. Heterochromatin plays a crucial role in driving the three-dimensional arrangement of the interphase genome, and in preserving genome stability by maintaining a subset of the genome in a silent state. Spatial genome organization contributes to normal patterns of gene function and expression, and is therefore of broad interest. Mammalian heterochromatin, the focus of this review, mainly localizes at the nuclear periphery, forming Lamina-associated domains (LADs), and at the nucleolar periphery, forming Nucleolus-associated domains (NADs). Together, these regions comprise approximately one-half of mammalian genomes, and most but not all loci within these domains are stochastically placed at either of these two locations after exit from mitosis at each cell cycle. Excitement about the role of these heterochromatic domains in early development has recently been heightened by the discovery that LADs appear at some loci in the preimplantation mouse embryo prior to other chromosomal features like compartmental identity and topologically-associated domains (TADs). While LADs have been extensively studied and mapped during cellular differentiation and early embryonic development, NADs have been less thoroughly studied. Here, we summarize pioneering studies of NADs and LADs, more recent advances in our understanding of cis/trans-acting factors that mediate these localizations, and discuss the functional significance of these associations.
Keywords: Chromosome; Development; Gene expression; Lamina-associated domain; Nucleolus-associated domain.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Heitz E, Das Heterochromatin der Moose., Jahrbücher Für Wissenschaftliche Bot. (1928).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
