

ATP13A3 is a major component of the enigmatic mammalian polyamine transport system
- PMID: 33310703
- PMCID: PMC7948421
- DOI: 10.1074/jbc.RA120.013908
ATP13A3 is a major component of the enigmatic mammalian polyamine transport system
Abstract
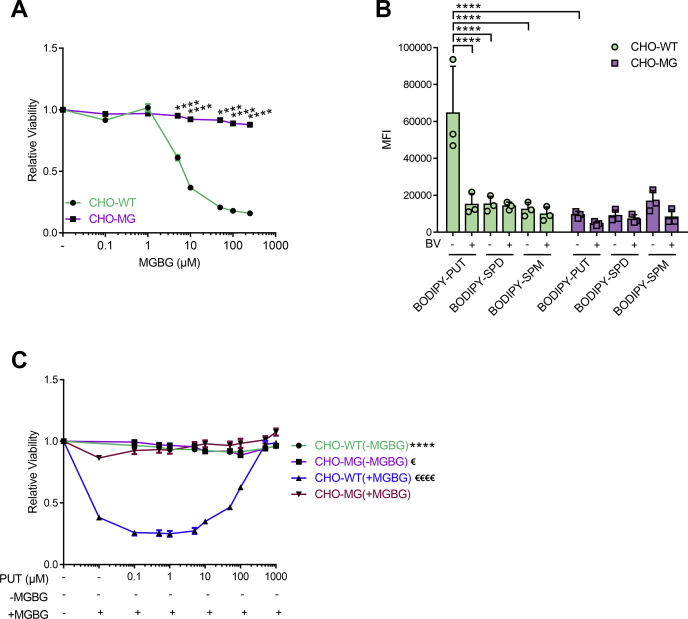
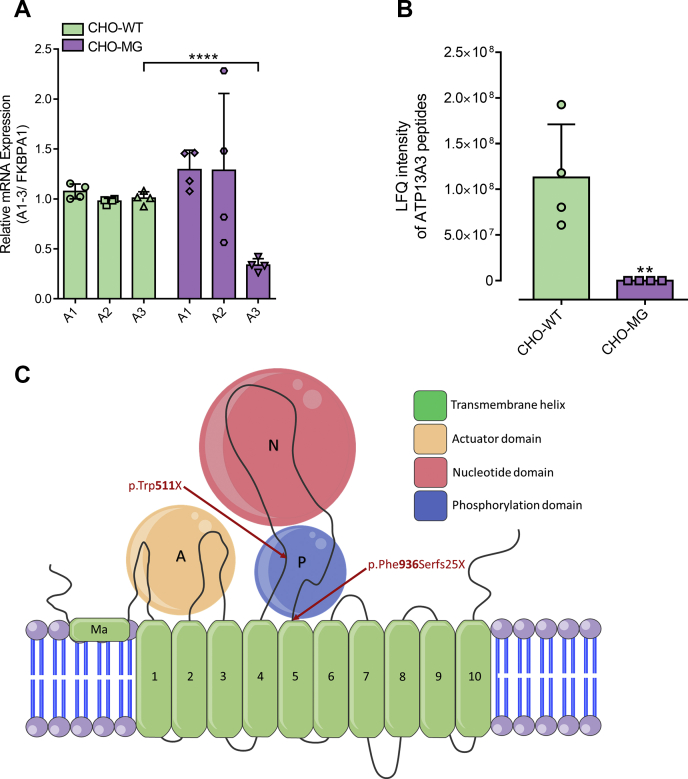
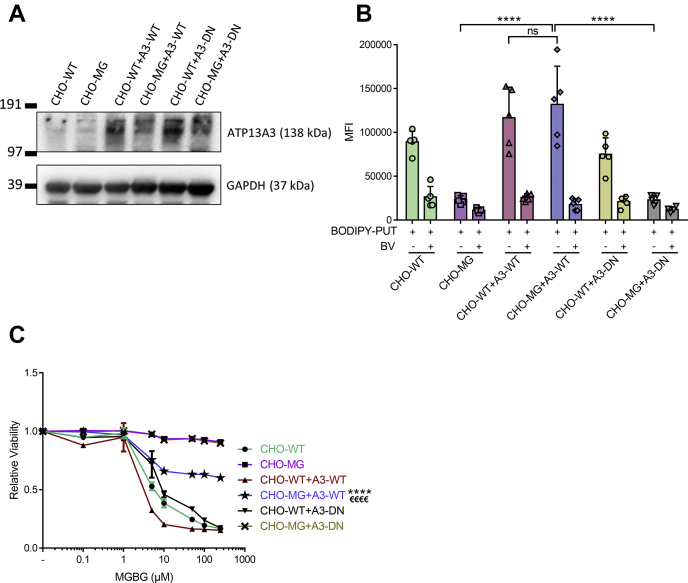
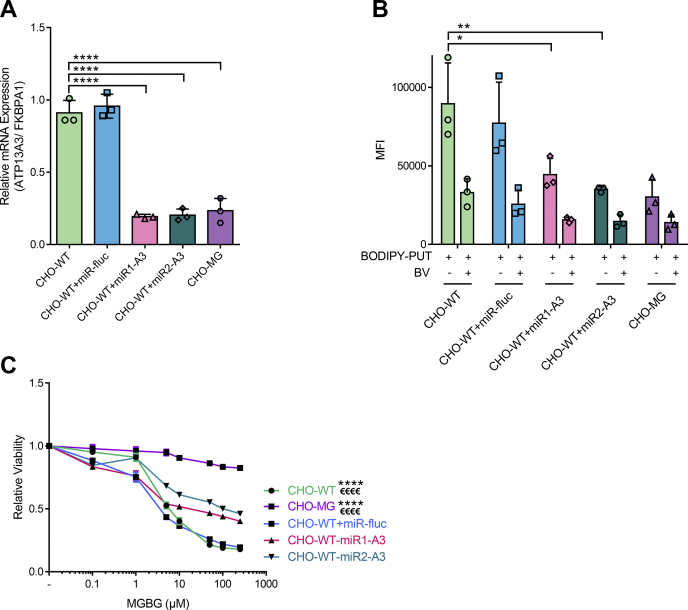
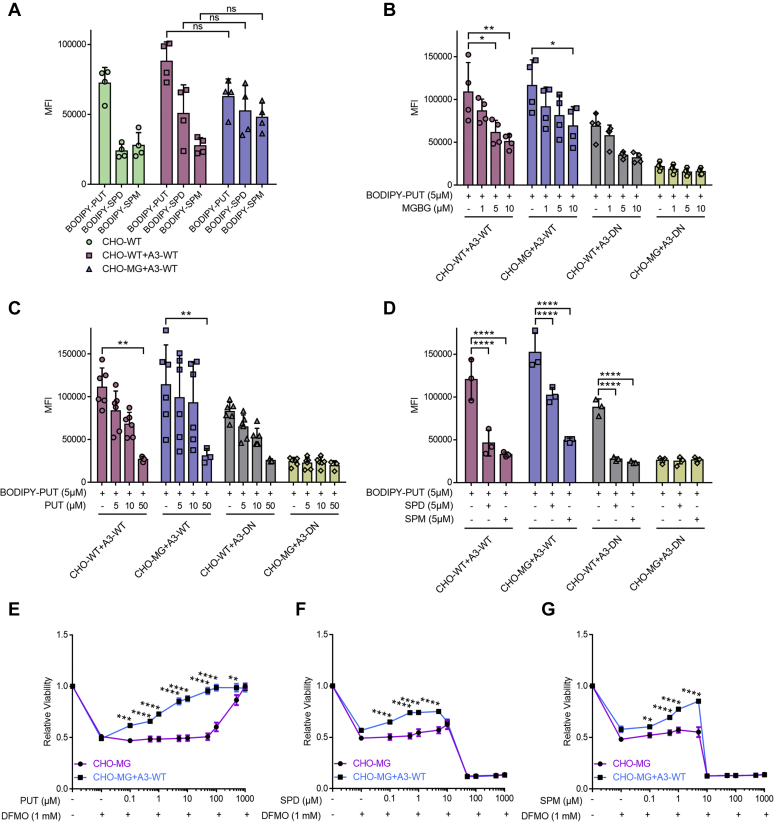
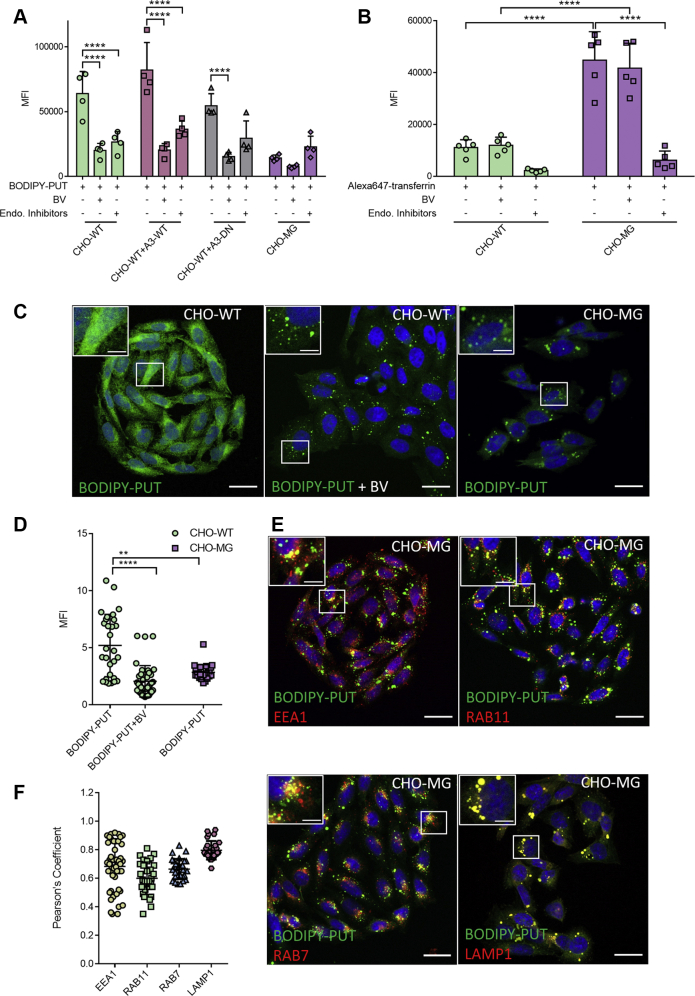
Polyamines, such as putrescine, spermidine, and spermine, are physiologically important polycations, but the transporters responsible for their uptake in mammalian cells remain poorly characterized. Here, we reveal a new component of the mammalian polyamine transport system using CHO-MG cells, a widely used model to study alternative polyamine uptake routes and characterize polyamine transport inhibitors for therapy. CHO-MG cells present polyamine uptake deficiency and resistance to a toxic polyamine biosynthesis inhibitor methylglyoxal bis-(guanylhydrazone) (MGBG), but the molecular defects responsible for these cellular characteristics remain unknown. By genome sequencing of CHO-MG cells, we identified mutations in an unexplored gene, ATP13A3, and found disturbed mRNA and protein expression. ATP13A3 encodes for an orphan P5B-ATPase (ATP13A3), a P-type transport ATPase that represents a candidate polyamine transporter. Interestingly, ATP13A3 complemented the putrescine transport deficiency and MGBG resistance of CHO-MG cells, whereas its knockdown in WT cells induced a CHO-MG phenotype demonstrated as a decrease in putrescine uptake and MGBG sensitivity. Taken together, our findings identify ATP13A3, which has been previously genetically linked with pulmonary arterial hypertension, as a major component of the mammalian polyamine transport system that confers sensitivity to MGBG.
Keywords: ATP13A3; P-type ATPase; P5B-ATPase; polyamine; polyamine transport system; putrescine; transporter.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Eisenberg T., Abdellatif M., Schroeder S., Primessnig U., Stekovic S., Pendl T., Harger A., Schipke J., Zimmermann A., Schmidt A., Tong M., Ruckenstuhl C., Dammbrueck C., Gross A.S., Herbst V. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat. Med. 2016;22:1428–1438. - PMC - PubMed
-
- Eisenberg T., Knauer H., Schauer A., Büttner S., Ruckenstuhl C., Carmona-Gutierrez D., Ring J., Schroeder S., Magnes C., Antonacci L., Fussi H., Deszcz L., Hartl R., Schraml E., Criollo A. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009;11:1305–1314. - PubMed
-
- Pegg A.E. Toxicity of polyamines and their metabolic products. Chem. Res. Toxicol. 2013;26:1782–1800. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
