

A dynamic, ring-forming MucB / RseB-like protein influences spore shape in Bacillus subtilis
- PMID: 33315869
- PMCID: PMC7769602
- DOI: 10.1371/journal.pgen.1009246
A dynamic, ring-forming MucB / RseB-like protein influences spore shape in Bacillus subtilis
Abstract
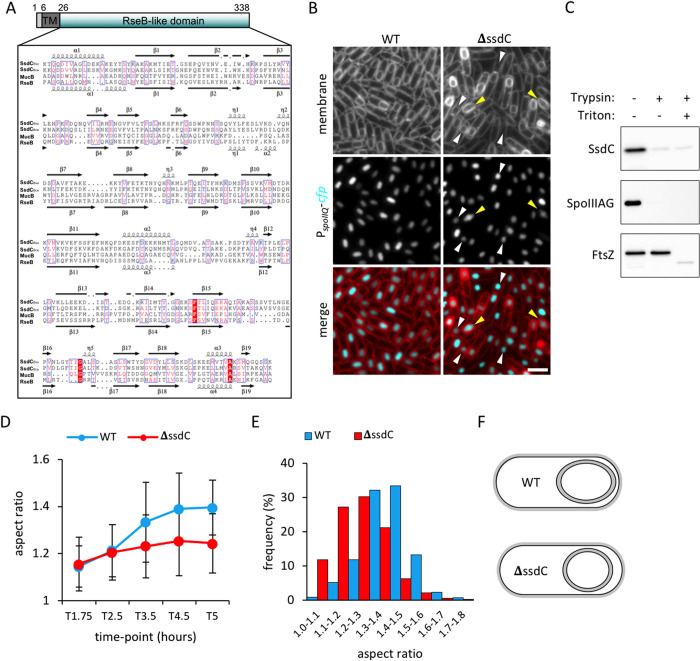
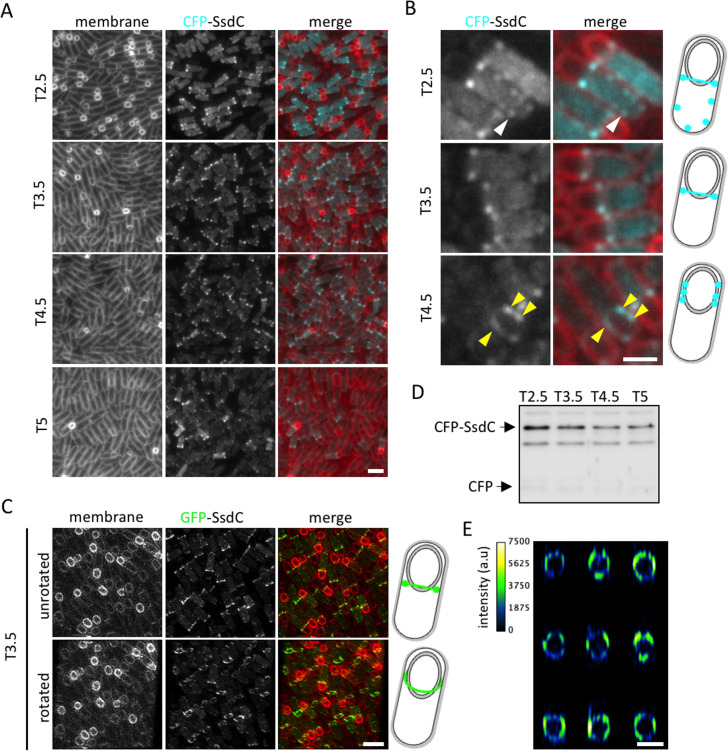
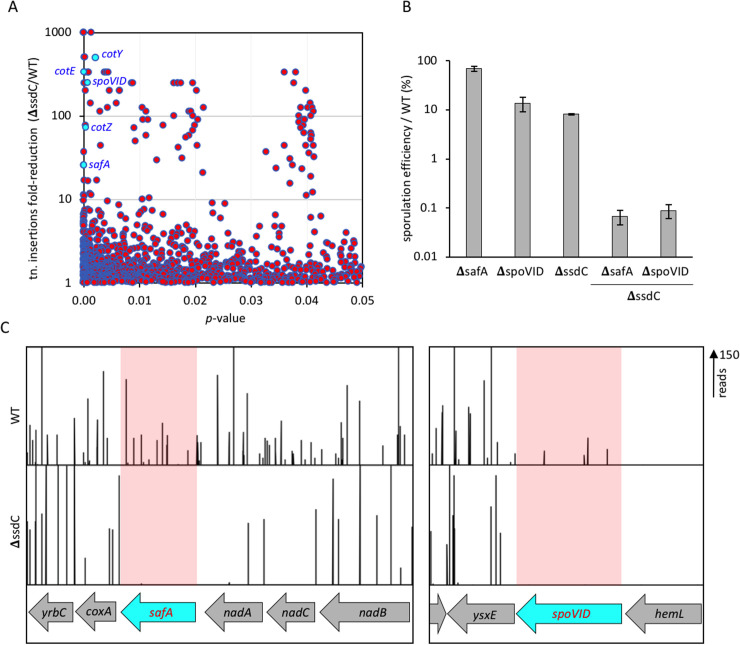
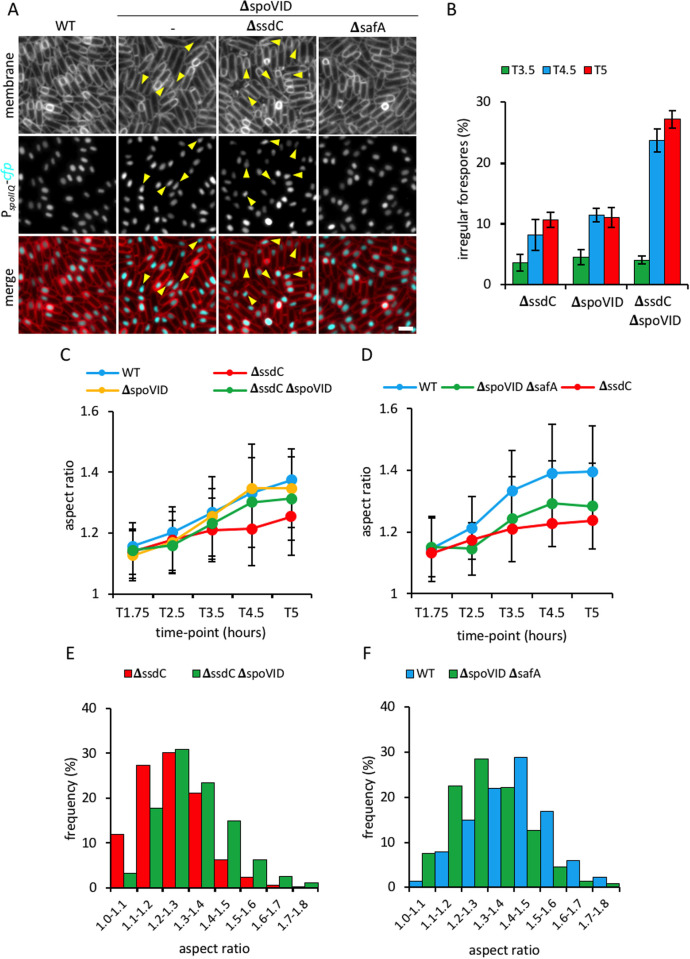
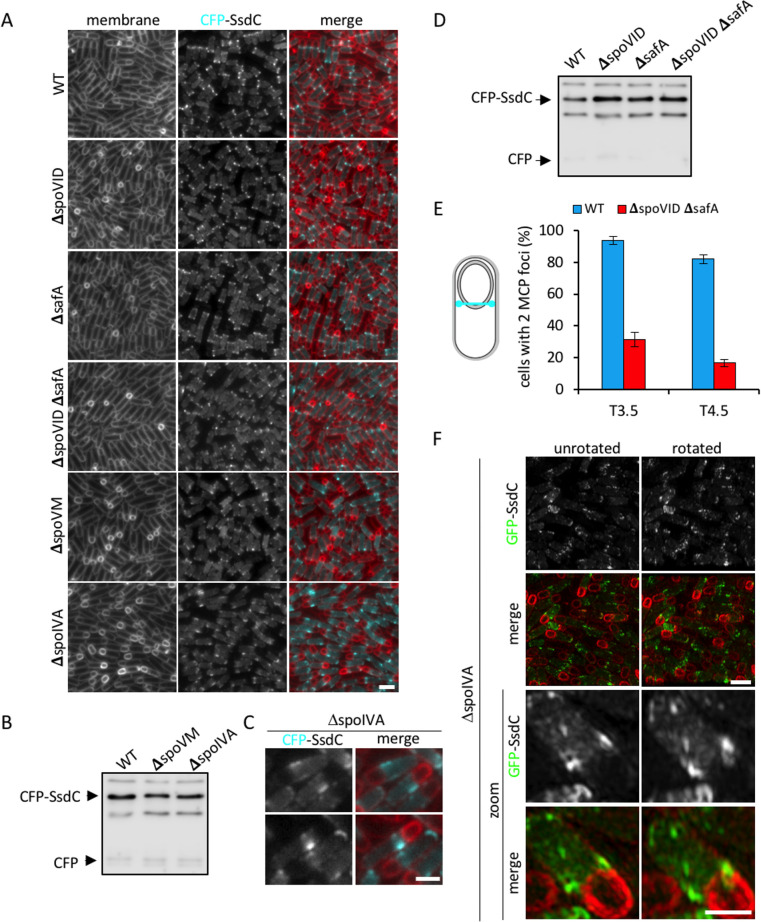
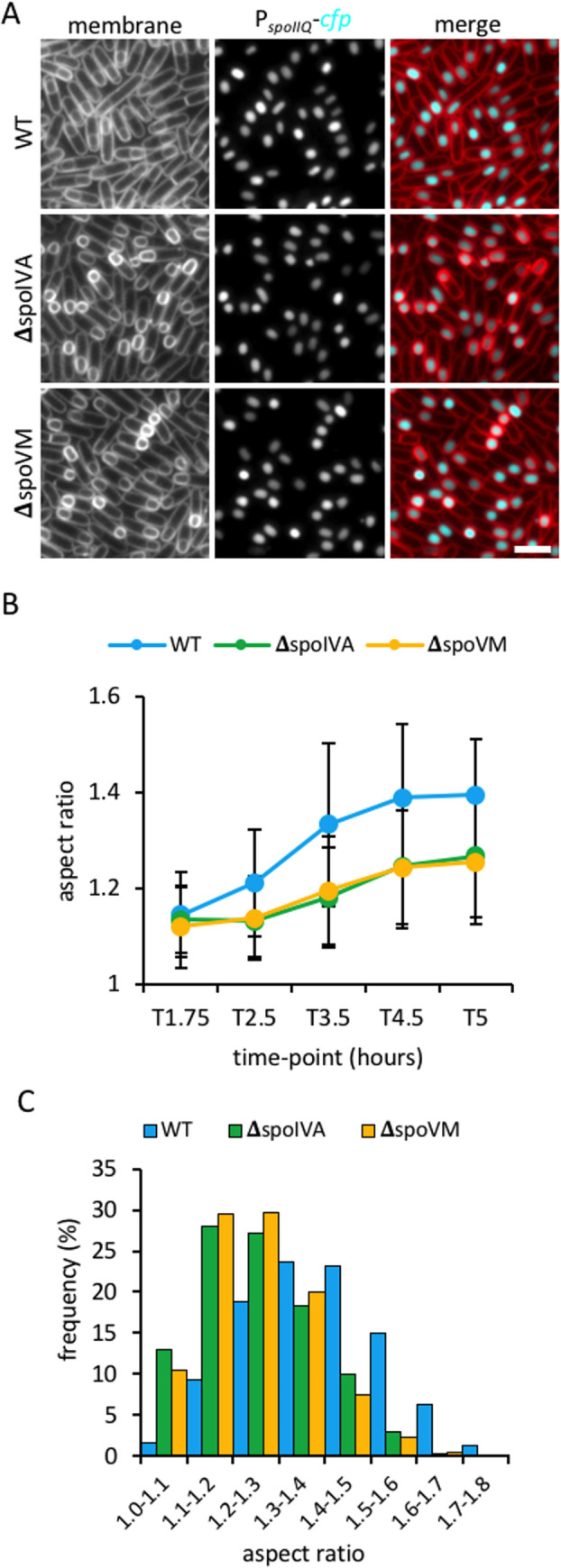
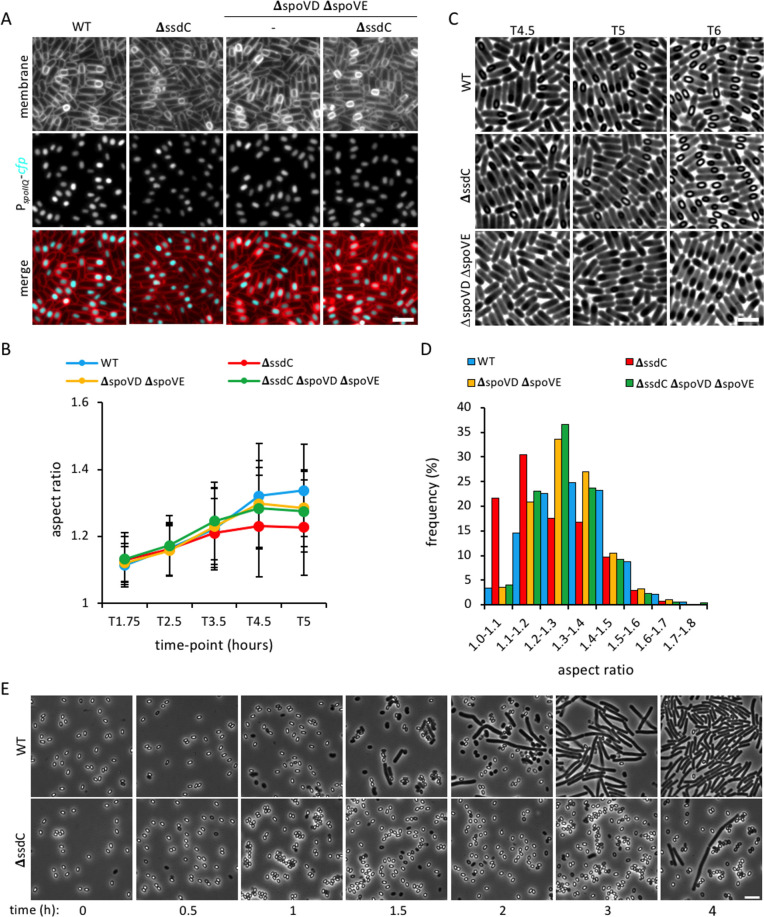
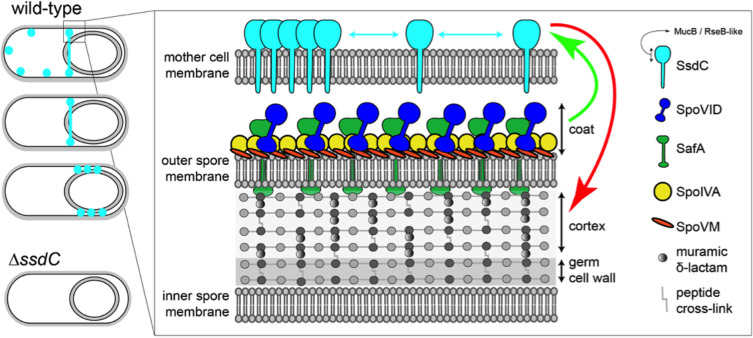
How organisms develop into specific shapes is a central question in biology. The maintenance of bacterial shape is connected to the assembly and remodelling of the cell envelope. In endospore-forming bacteria, the pre-spore compartment (the forespore) undergoes morphological changes that result in a spore of defined shape, with a complex, multi-layered cell envelope. However, the mechanisms that govern spore shape remain poorly understood. Here, using a combination of fluorescence microscopy, quantitative image analysis, molecular genetics and transmission electron microscopy, we show that SsdC (formerly YdcC), a poorly-characterized new member of the MucB / RseB family of proteins that bind lipopolysaccharide in diderm bacteria, influences spore shape in the monoderm Bacillus subtilis. Sporulating cells lacking SsdC fail to adopt the typical oblong shape of wild-type forespores and are instead rounder. 2D and 3D-fluorescence microscopy suggest that SsdC forms a discontinuous, dynamic ring-like structure in the peripheral membrane of the mother cell, near the mother cell proximal pole of the forespore. A synthetic sporulation screen identified genetic relationships between ssdC and genes involved in the assembly of the spore coat. Phenotypic characterization of these mutants revealed that spore shape, and SsdC localization, depend on the coat basement layer proteins SpoVM and SpoIVA, the encasement protein SpoVID and the inner coat protein SafA. Importantly, we found that the ΔssdC mutant produces spores with an abnormal-looking cortex, and abolishing cortex synthesis in the mutant largely suppresses its shape defects. Thus, SsdC appears to play a role in the proper assembly of the spore cortex, through connections to the spore coat. Collectively, our data suggest functional diversification of the MucB / RseB protein domain between diderm and monoderm bacteria and identify SsdC as an important factor in spore shape development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
