

Active bacterial modification of the host environment through RNA polymerase II inhibition
- PMID: 33320835
- PMCID: PMC7880420
- DOI: 10.1172/JCI140333
Active bacterial modification of the host environment through RNA polymerase II inhibition
Abstract
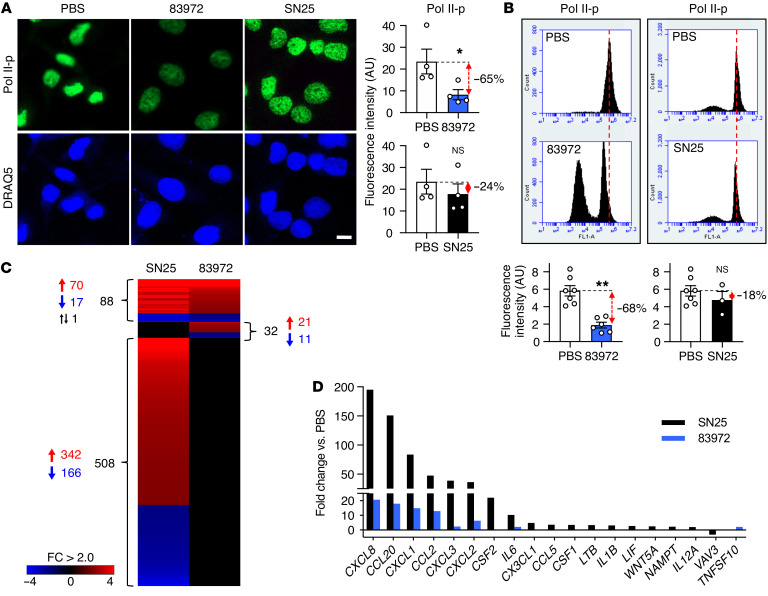
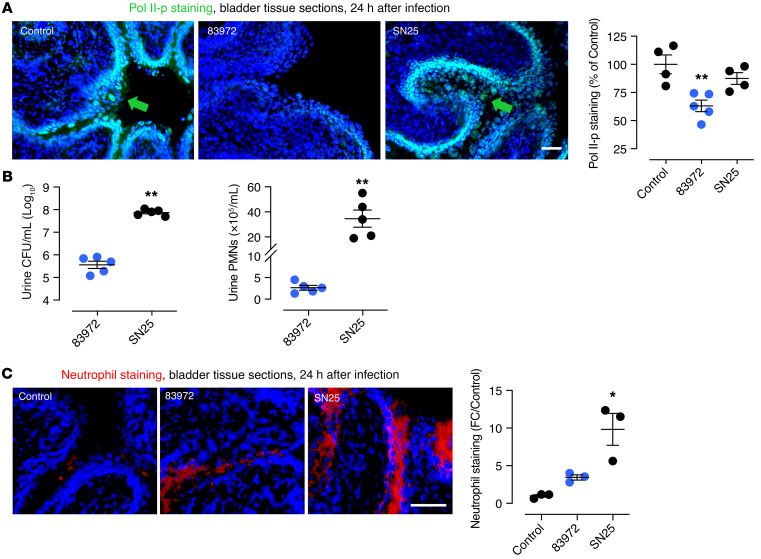
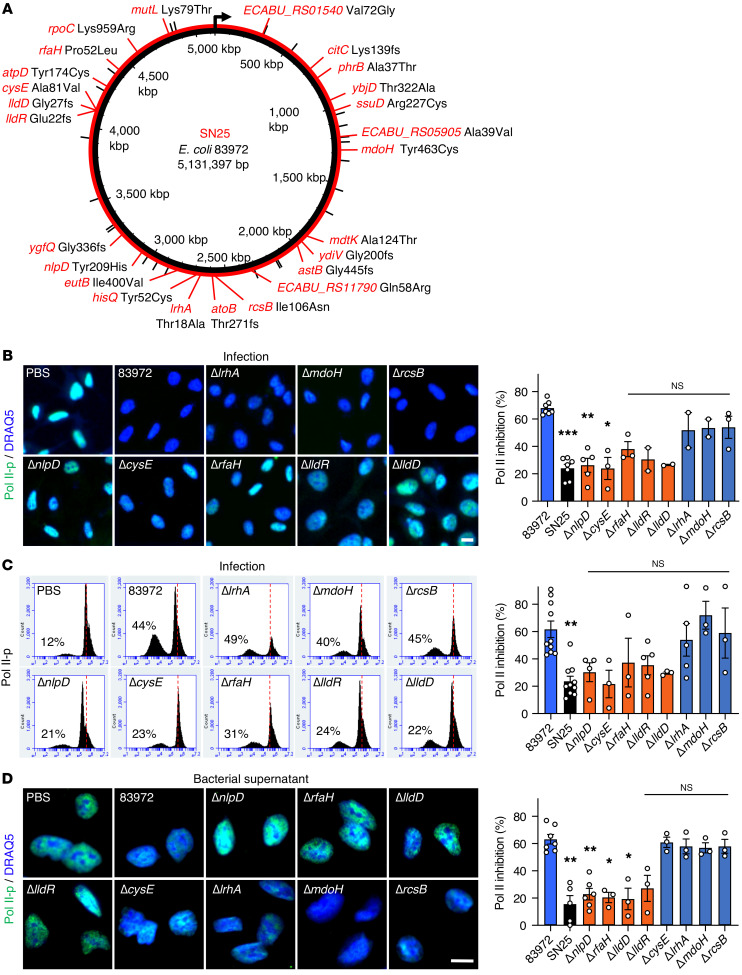
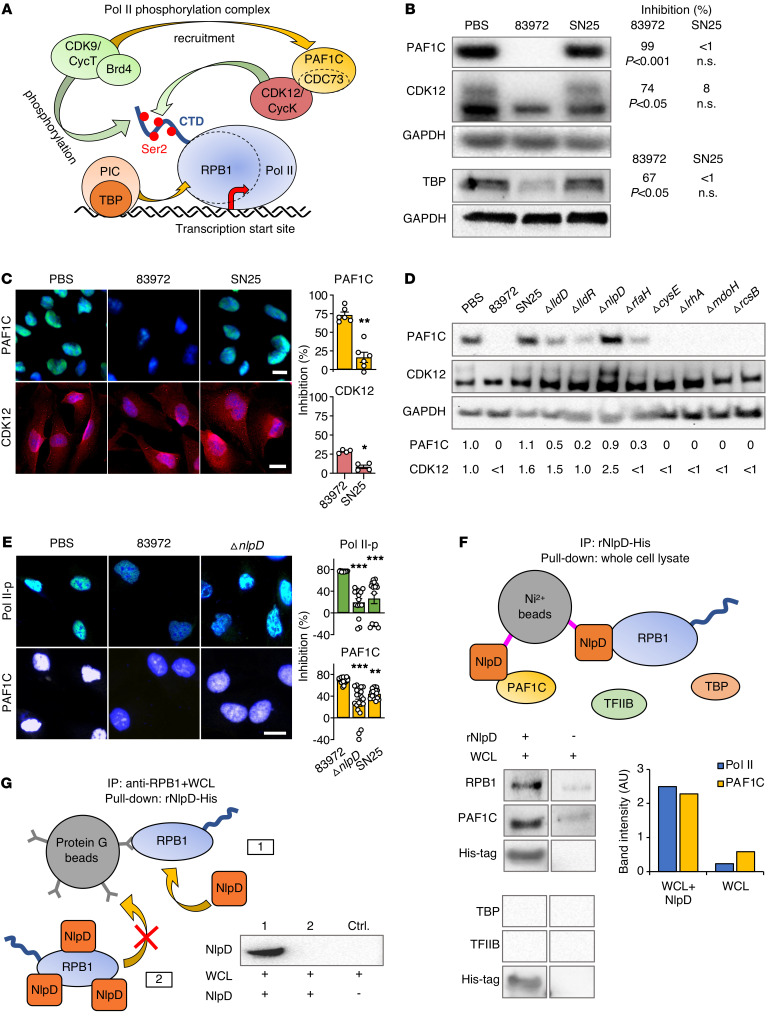
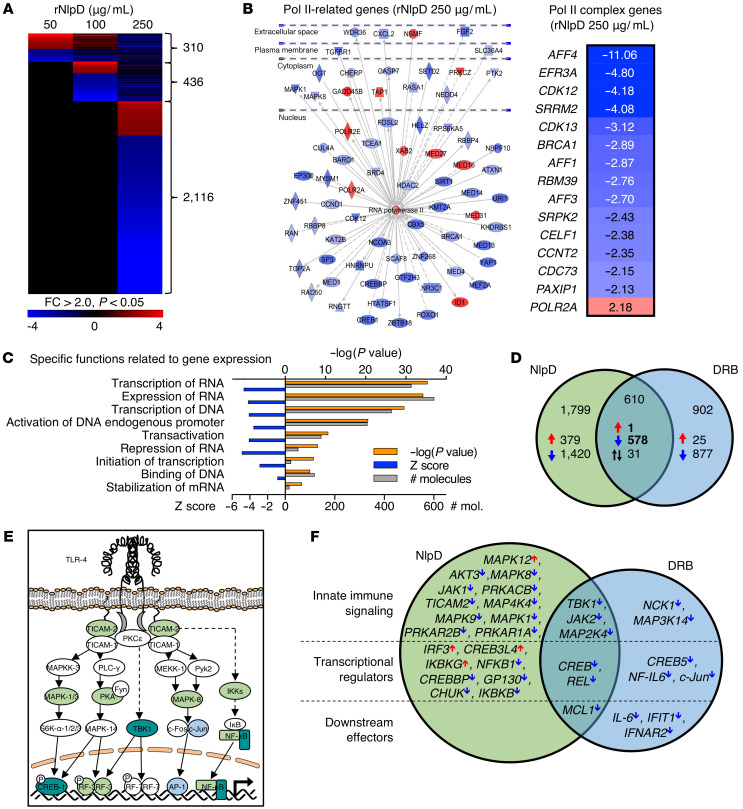
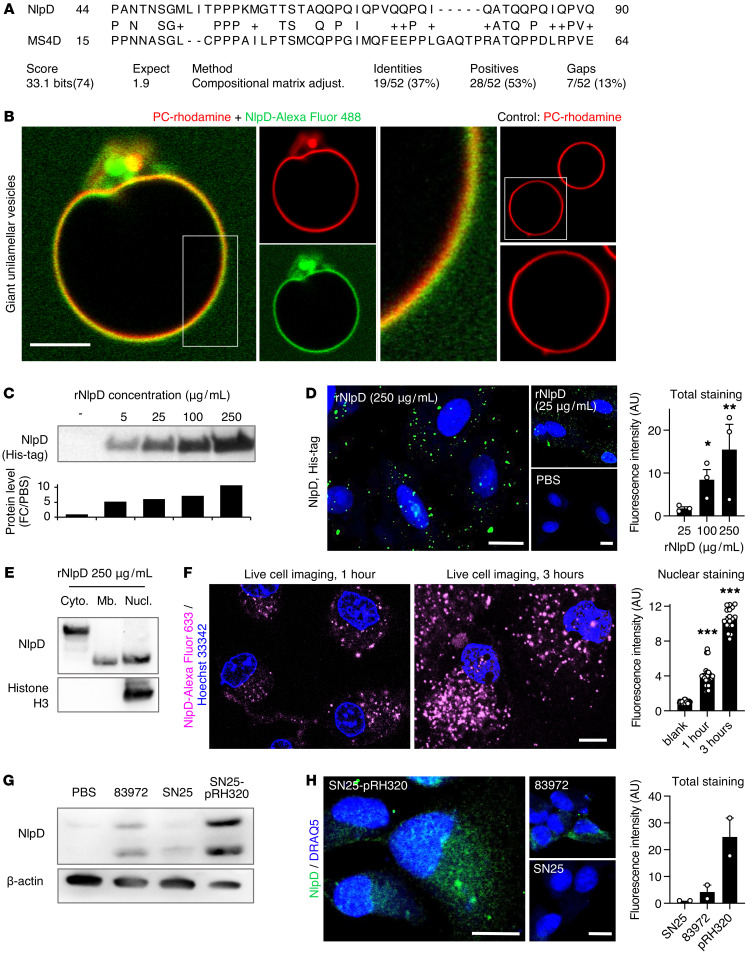
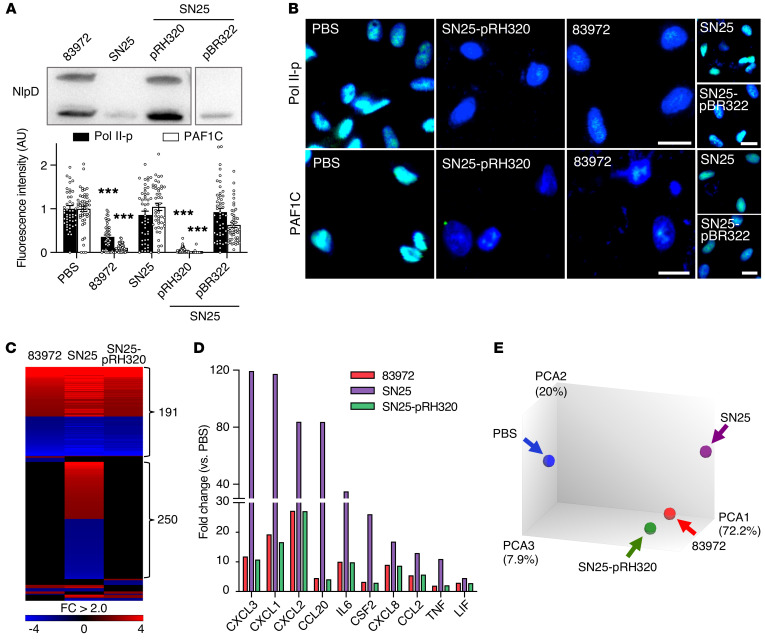
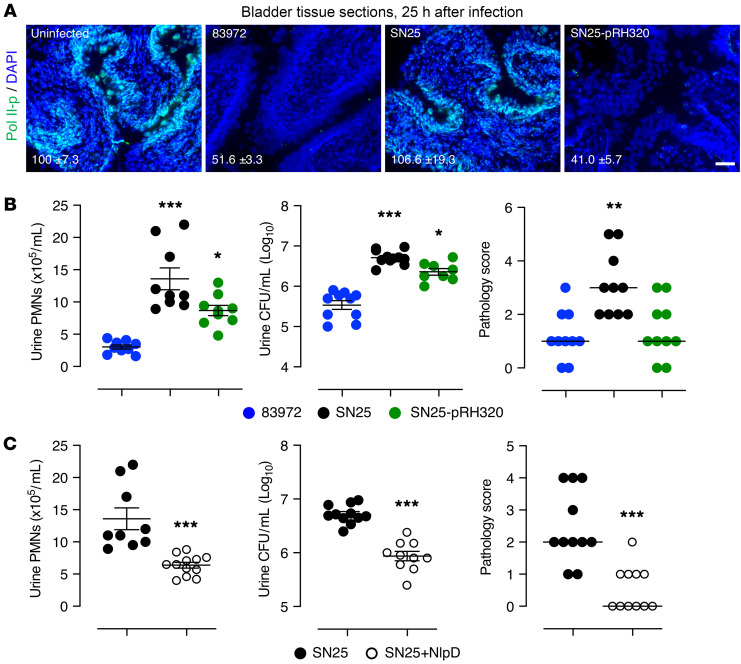
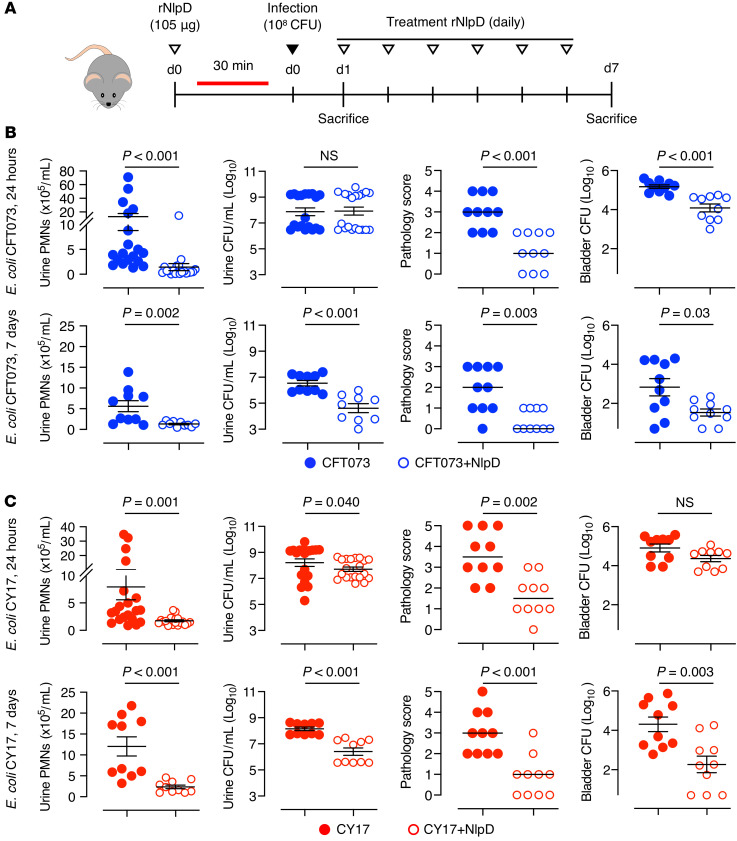
Unlike pathogens, which attack the host, commensal bacteria create a state of friendly coexistence. Here, we identified a mechanism of bacterial adaptation to the host niche, where they reside. Asymptomatic carrier strains were shown to inhibit RNA polymerase II (Pol II) in host cells by targeting Ser2 phosphorylation, a step required for productive mRNA elongation. Assisted by a rare, spontaneous loss-of-function mutant from a human carrier, the bacterial NlpD protein was identified as a Pol II inhibitor. After internalization by host cells, NlpD was shown to target constituents of the Pol II phosphorylation complex (RPB1 and PAF1C), attenuating host gene expression. Therapeutic efficacy of a recombinant NlpD protein was demonstrated in a urinary tract infection model, by reduced tissue pathology, accelerated bacterial clearance, and attenuated Pol II-dependent gene expression. The findings suggest an intriguing, evolutionarily conserved mechanism for bacterial modulation of host gene expression, with a remarkable therapeutic potential.
Keywords: Immunotherapy; Inflammation; Microbiology; Transcription.
Conflict of interest statement
Figures
Comment in
-
Collateral effects of deletion of nlpD on rpoS and rpoS-dependent genes.J Clin Invest. 2021 Sep 15;131(18):e152693. doi: 10.1172/JCI152693. J Clin Invest. 2021. PMID: 34523610 Free PMC article. No abstract available.
-
Collateral effects of deletion of nlpD on rpoS and rpoS-dependent genes. Reply.J Clin Invest. 2021 Sep 15;131(18):e153234. doi: 10.1172/JCI153234. J Clin Invest. 2021. PMID: 34523612 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
