

α-Crystallins in the Vertebrate Eye Lens: Complex Oligomers and Molecular Chaperones
- PMID: 33321054
- PMCID: PMC8062273
- DOI: 10.1146/annurev-physchem-090419-121428
α-Crystallins in the Vertebrate Eye Lens: Complex Oligomers and Molecular Chaperones
Abstract
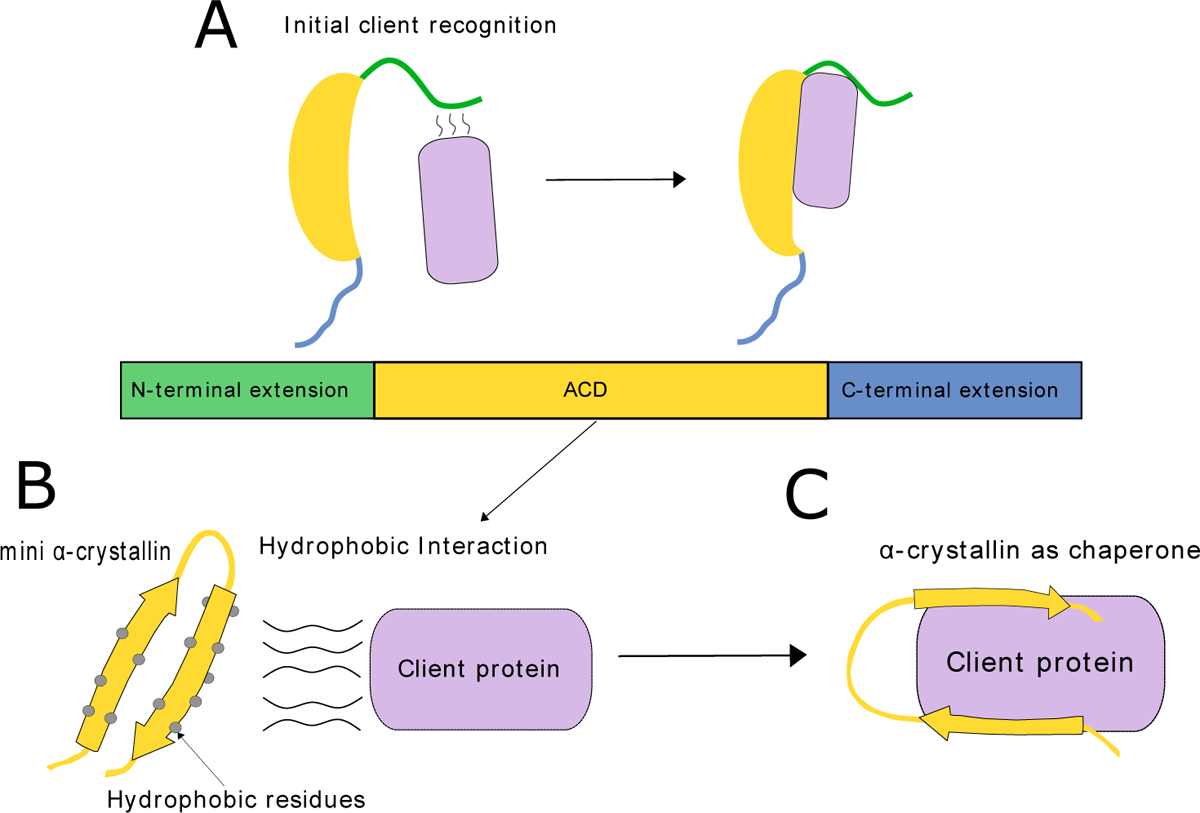
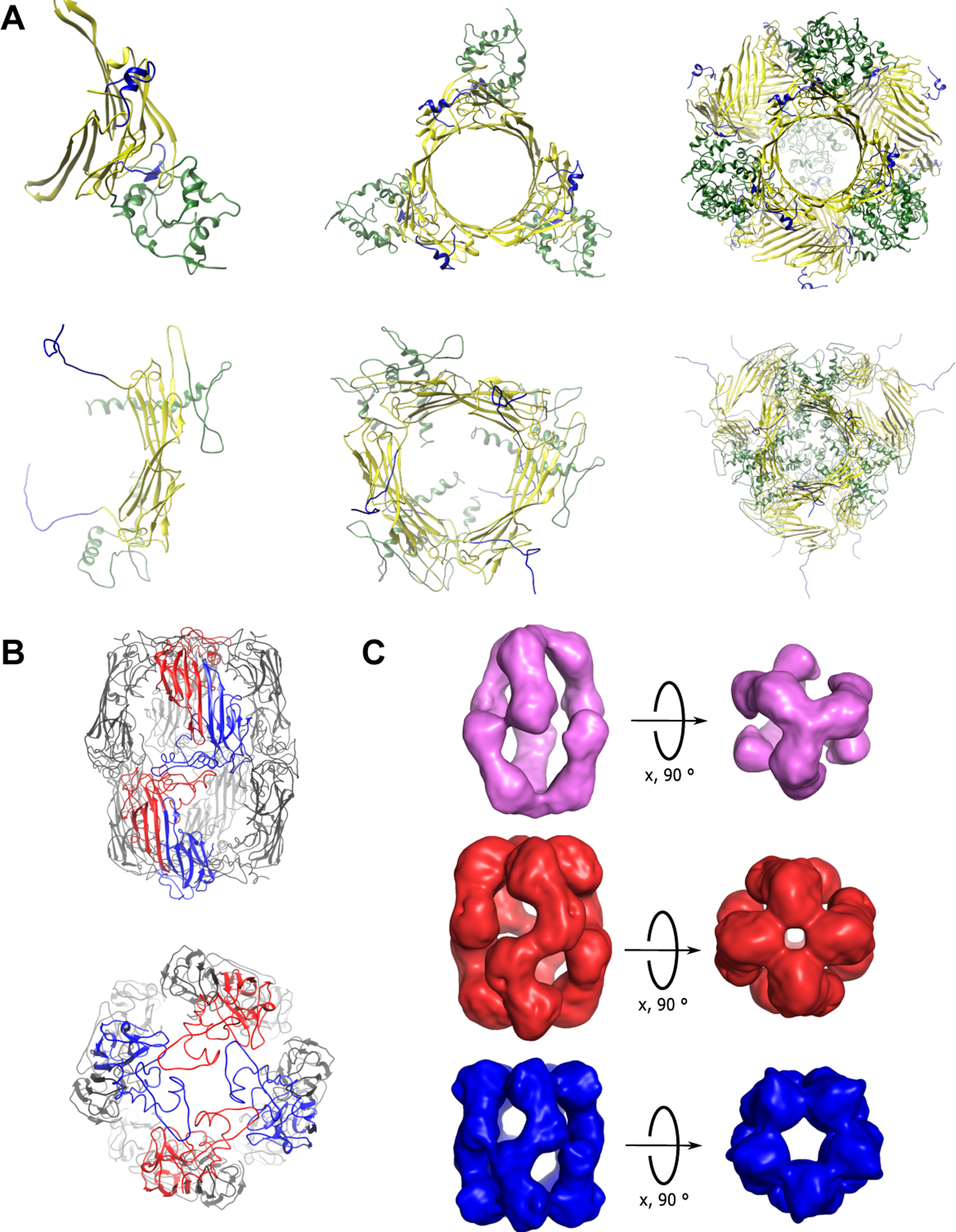
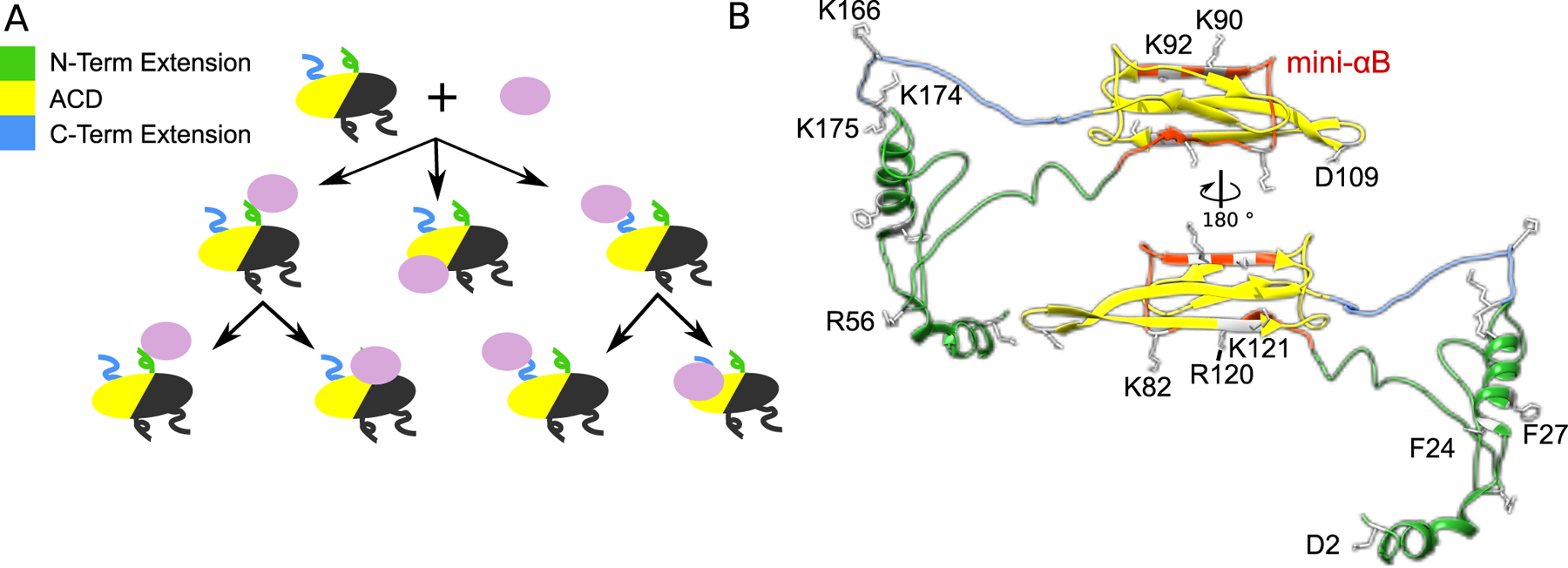
α-Crystallins are small heat-shock proteins that act as holdase chaperones. In humans, αA-crystallin is expressed only in the eye lens, while αB-crystallin is found in many tissues. α-Crystallins have a central domain flanked by flexible extensions and form dynamic, heterogeneous oligomers. Structural models show that both the C- and N-terminal extensions are important for controlling oligomerization through domain swapping. α-Crystallin prevents aggregation of damaged β- and γ-crystallins by binding to the client protein using a variety of binding modes. α-Crystallin chaperone activity can be compromised by mutation or posttranslational modifications, leading to protein aggregation and cataract. Because of their high solubility and their ability to form large, functional oligomers, α-crystallins are particularly amenable to structure determination by solid-state nuclear magnetic resonance (NMR) and solution NMR, as well as cryo-electron microscopy.
Keywords: intermolecular interactions; molecular chaperone; protein oligomer; protein solubility; vertebrate lens protein; α-crystallin.
Figures
References
-
- Dilley Keith J. and Harding John J.. Changes in proteins of the human lens in development and aging. Biochimica et Biophysica Acta (BBA) - Protein Structure, 386(2):391–408, April 1975. ISSN 0005–2795.. - PubMed
-
- Delaye Mireille and Tardieu Annette. Short-range order of crystallin proteins accounts for eye lens transparency. Nature, 302(5907):302415a0, March 1983. ISSN 1476–4687.. - PubMed
-
- Kröger Ronald H. H., Campbell Melanie C. W., Munger Rejean, and Fernald Russell D.. Refractive index distribution and spherical aberration in the crystalline lens of the African cichlid fish Haplochromis burtoni. Vision Research, 34(14):1815–1822, 1994. - PubMed
