

Reductive TCA cycle metabolism fuels glutamine- and glucose-stimulated insulin secretion
- PMID: 33321098
- PMCID: PMC8115731
- DOI: 10.1016/j.cmet.2020.11.020
Reductive TCA cycle metabolism fuels glutamine- and glucose-stimulated insulin secretion
Abstract
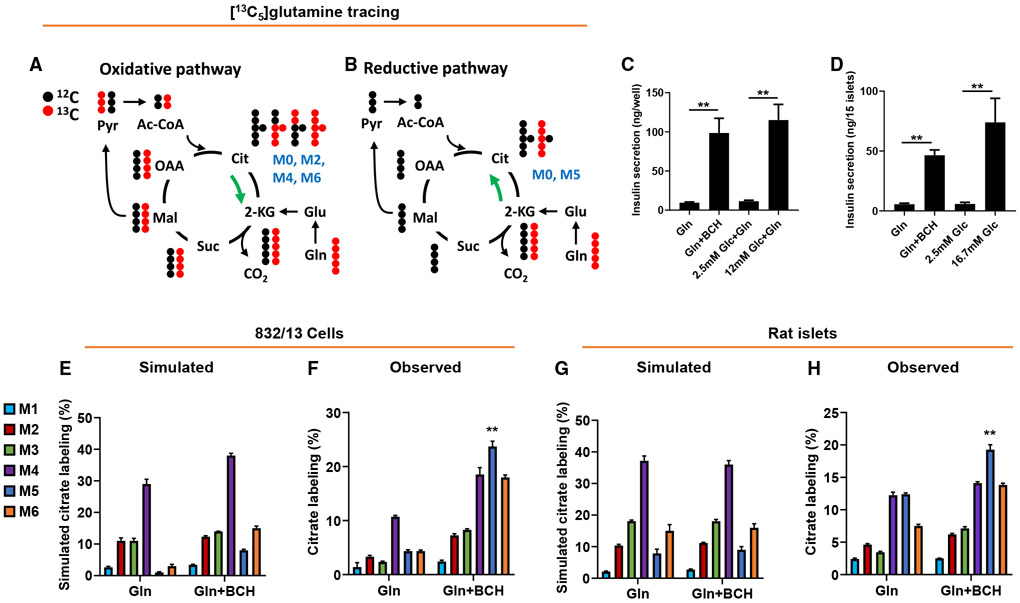
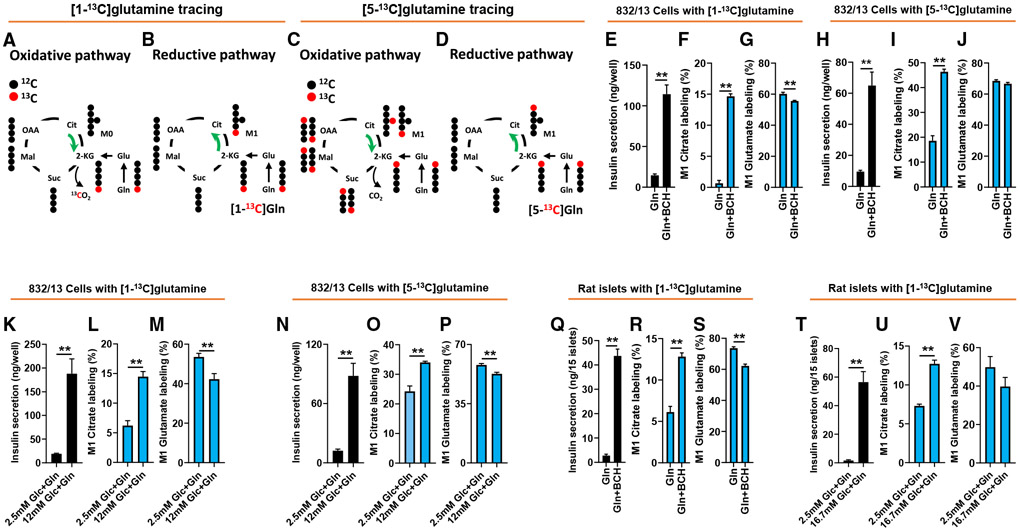
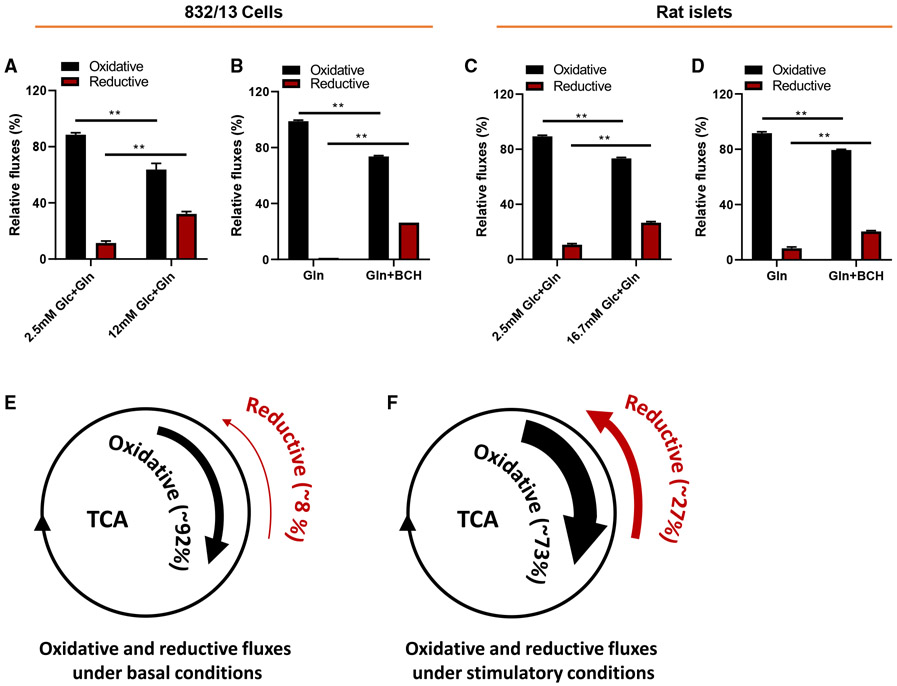
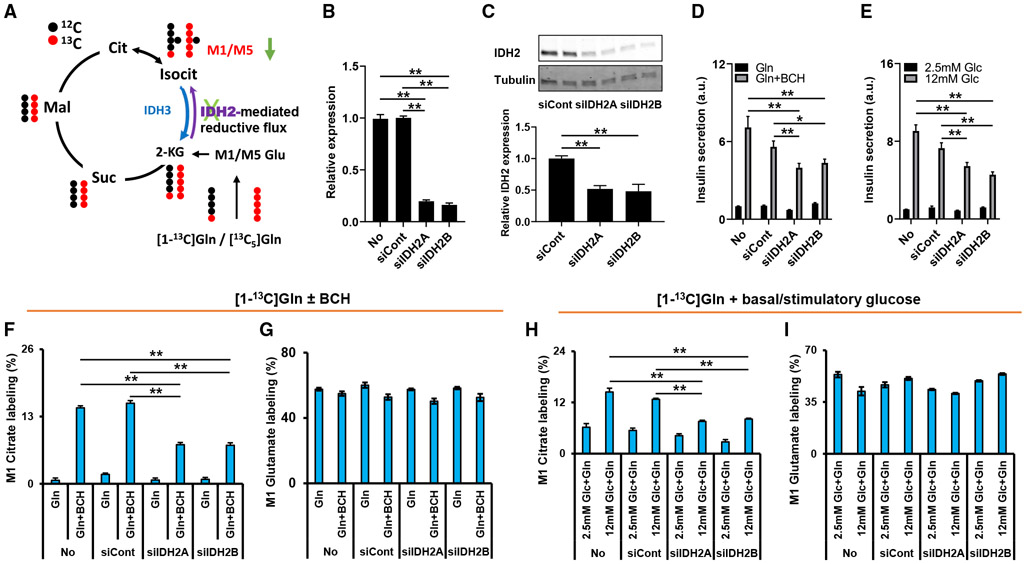
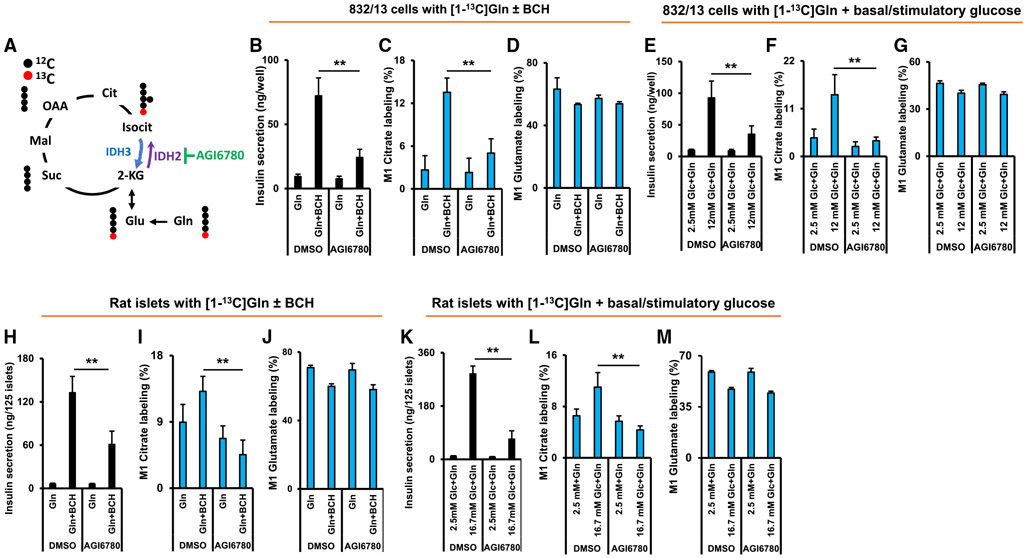
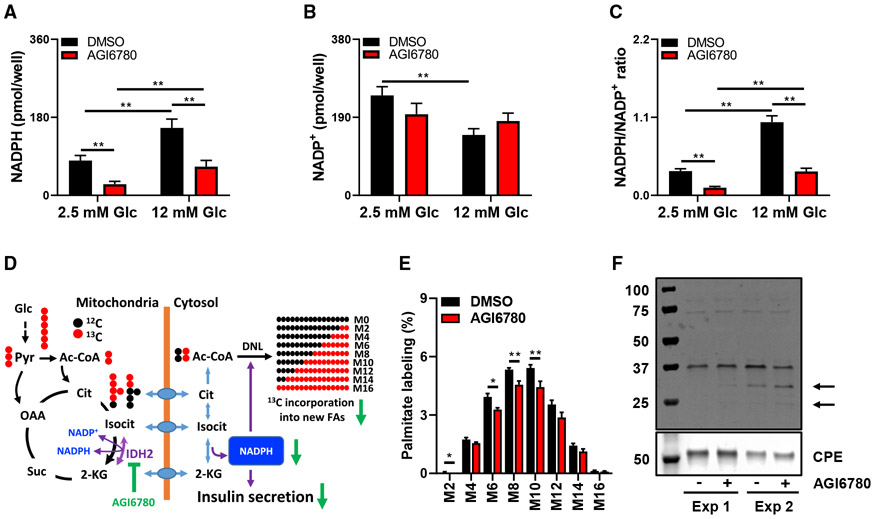
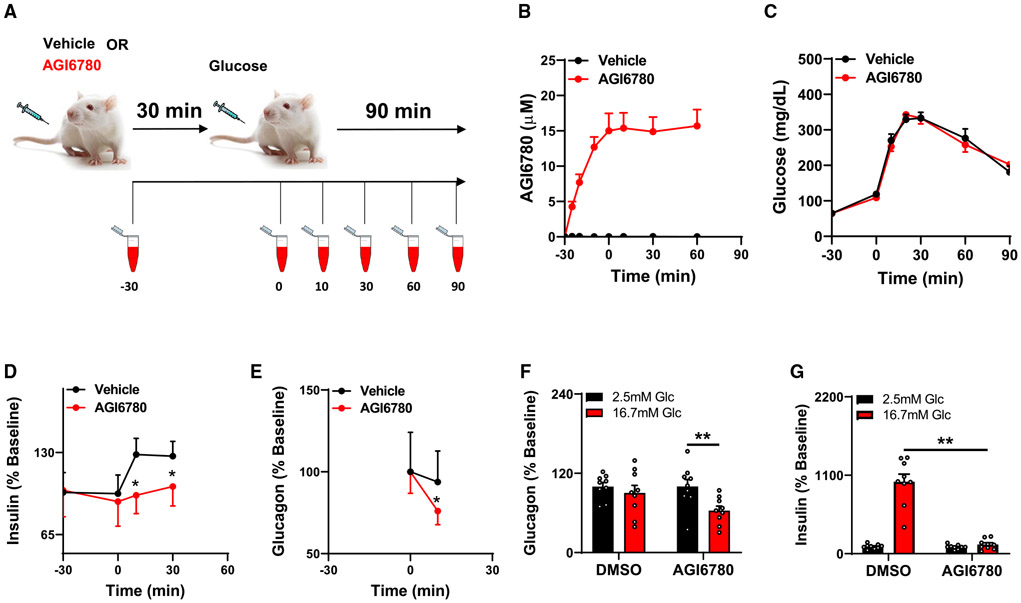
Metabolic fuels regulate insulin secretion by generating second messengers that drive insulin granule exocytosis, but the biochemical pathways involved are incompletely understood. Here we demonstrate that stimulation of rat insulinoma cells or primary rat islets with glucose or glutamine + 2-aminobicyclo-(2,2,1)-heptane-2-carboxylic acid (Gln + BCH) induces reductive, "counter-clockwise" tricarboxylic acid (TCA) cycle flux of glutamine to citrate. Molecular or pharmacologic suppression of isocitrate dehydrogenase-2 (IDH2), which catalyzes reductive carboxylation of 2-ketoglutarate to isocitrate, results in impairment of glucose- and Gln + BCH-stimulated reductive TCA cycle flux, lowering of NADPH levels, and inhibition of insulin secretion. Pharmacologic suppression of IDH2 also inhibits insulin secretion in living mice. Reductive TCA cycle flux has been proposed as a mechanism for generation of biomass in cancer cells. Here we demonstrate that reductive TCA cycle flux also produces stimulus-secretion coupling factors that regulate insulin secretion, including in non-dividing cells.
Keywords: NADPH; anaplerosis; insulin secretion; isocitrate dehydrogenase-2; metabolic flux; pancreatic islet β cells; reductive TCA cycle; stable isotopes.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest in conduct of this research. C.B.N. is a paid consultant for Eli Lilly, Axcella Health, Boehringer Ingelheim, and Sigilon. Whereas all of these companies have interests in diabetes therapy, they have no involvement or competing interests in the research described in this paper.
Figures
Comment in
-
Pancreatic β cells put the glutamine engine in reverse.Cell Metab. 2021 Apr 6;33(4):702-704. doi: 10.1016/j.cmet.2021.03.010. Cell Metab. 2021. PMID: 33826912
References
-
- Ashcroft FM, Harrison DE, and Ashcroft SJ (1984). Glucose induces closure of single potassium channels in isolated rat pancreatic beta-cells. Nature 312, 446–448. - PubMed
-
- Cook DL, and Hales CN (1984). Intracellular ATP directly blocks K+ channels in pancreatic B-cells. Nature 311, 271–273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
