

Rolling-translated EGFR variants sustain EGFR signaling and promote glioblastoma tumorigenicity
- PMID: 33325513
- PMCID: PMC8099477
- DOI: 10.1093/neuonc/noaa279
Rolling-translated EGFR variants sustain EGFR signaling and promote glioblastoma tumorigenicity
Abstract
Background: Aberrant epidermal growth factor receptor (EGFR) activation is observed in over 50% of cases of adult glioblastoma (GBM). Nevertheless, EGFR antibodies are ineffective in clinical GBM treatment, suggesting the existence of redundant EGFR activation mechanisms. Whether circular RNA (circRNA) encodes a protein involved in EGFR-driven GBM remains unclear. We reported an unexpected mechanism in which circular EGFR RNA (circ-EGFR) encodes a novel EGFR variant to sustained EGFR activation.
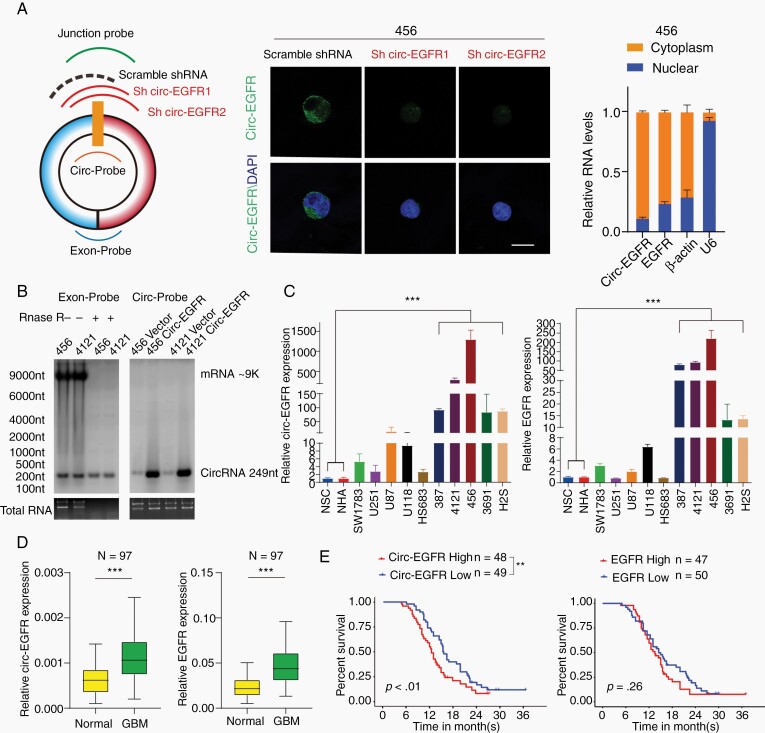
Method: We used RNA-seq, Northern blot, and Sanger sequencing to confirm the existence of circ-EGFR. Antibodies and a liquid chromatograph tandem mass spectrometer were used to identify circ-EGFR protein products. Lentivirus-transfected stable cell lines were used to assess the biological functions of the novel protein in vitro and in vivo. Clinical implications of circ-EGFR were assessed using 97 pathologically diagnosed GBM patient samples.
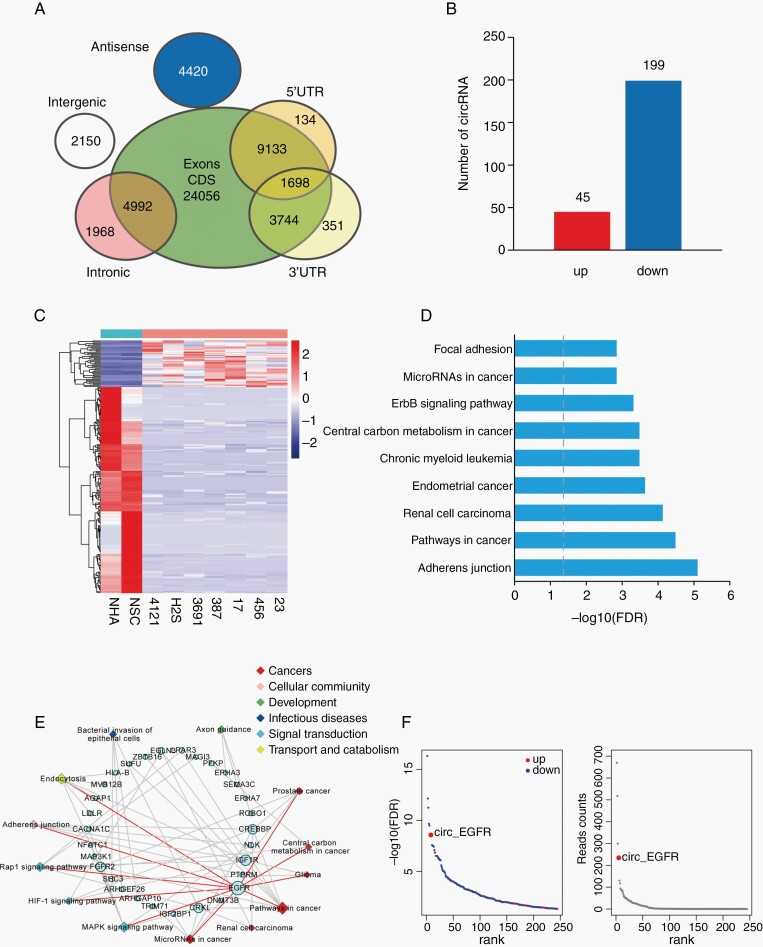
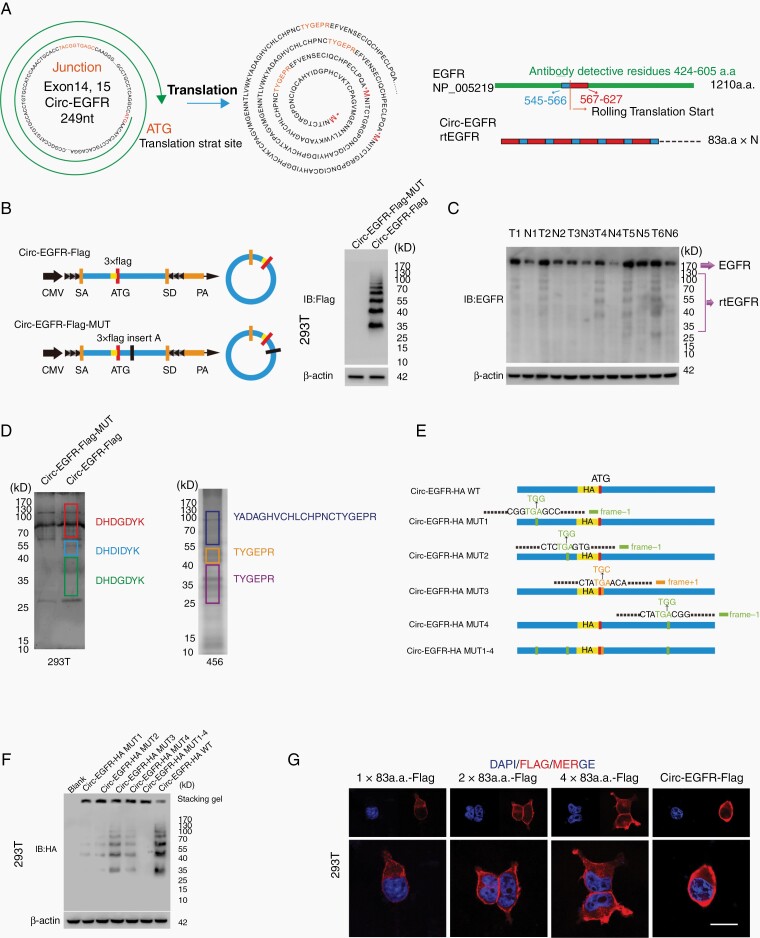
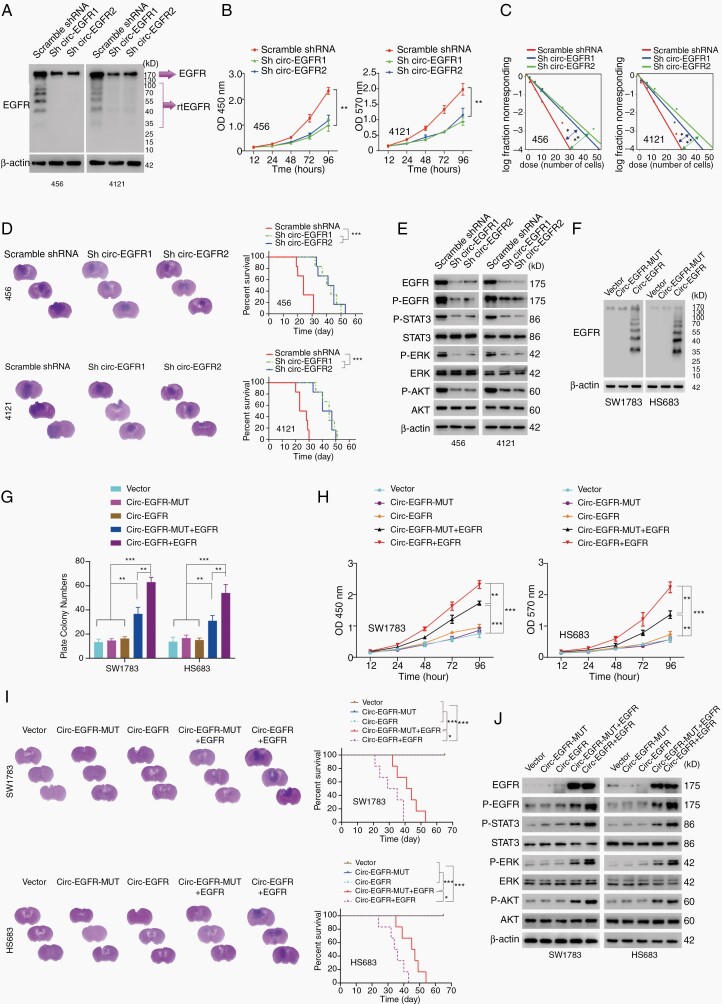
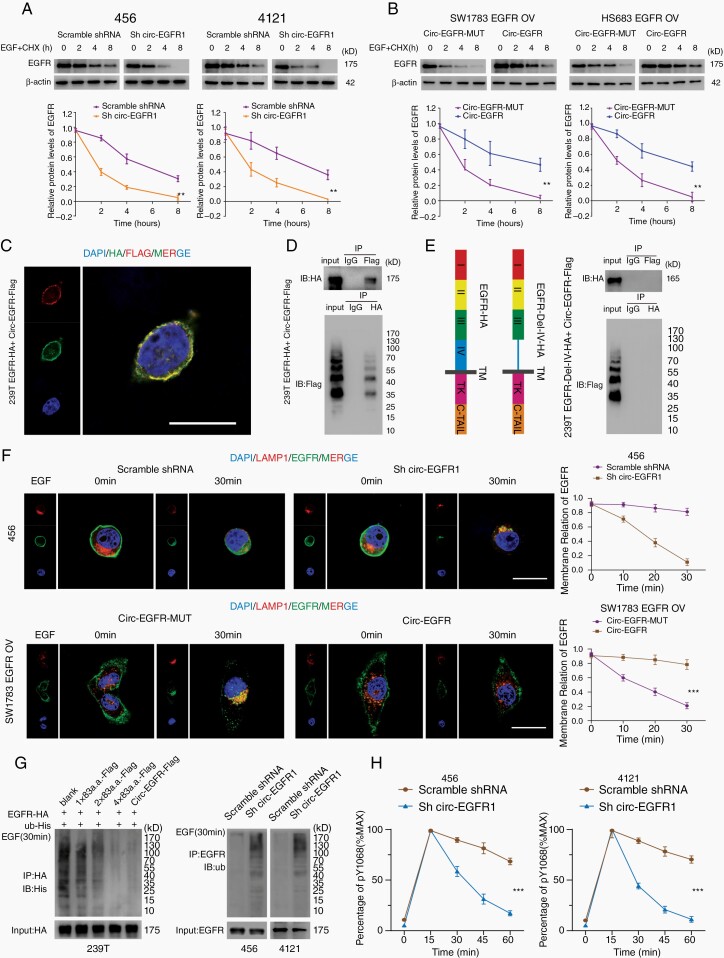
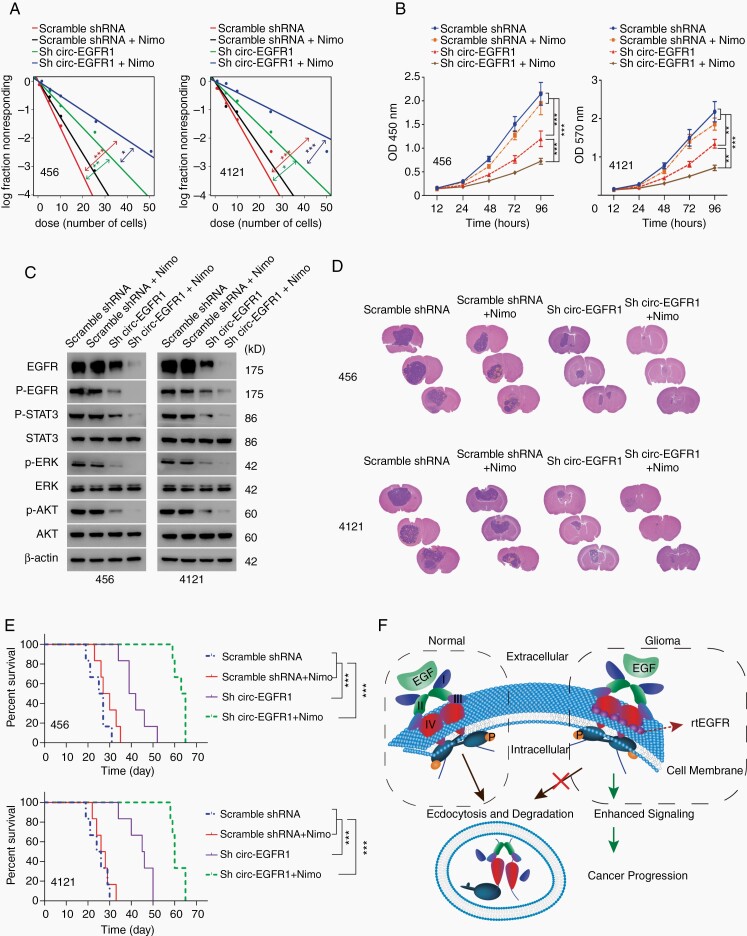
Results: The infinite open reading frame (iORF) in circ-EGFR translated repeating amino acid sequences via rolling translation and programmed -1 ribosomal frameshifting (-1PRF) induced out-of-frame stop codon (OSC), forming a polymetric novel protein-complex, which we termed rolling-translated EGFR (rtEGFR). rtEGFR directly interacted with EGFR, maintained EGFR membrane localization and attenuated EGFR endocytosis and degradation. Importantly, circ-EGFR levels correlated with the EGFR signature and predicted the poor prognosis of GBM patients. Deprivation of rtEGFR in brain tumor-initiating cells (BTICs) attenuated tumorigenicity and enhanced the anti-GBM effect.
Conclusion: Our findings identified the endogenous rolling-translated protein and provided strong clinical evidence that targeting rtEGFR could improve the efficiency of EGFR-targeting therapies in GBM.
Keywords: CircRNA; EGFR; glioblastoma; iORF; rolling translation.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Comment in
-
You spin me right 'round.Neuro Oncol. 2021 May 5;23(5):707-708. doi: 10.1093/neuonc/noab032. Neuro Oncol. 2021. PMID: 33704479 Free PMC article. No abstract available.
References
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
