

Self-assembly-based posttranslational protein oscillators
- PMID: 33328225
- PMCID: PMC7744077
- DOI: 10.1126/sciadv.abc1939
Self-assembly-based posttranslational protein oscillators
Abstract
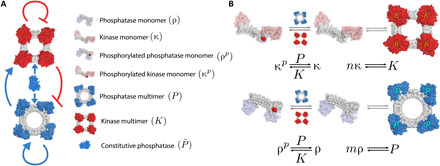
Recent advances in synthetic posttranslational protein circuits are substantially impacting the landscape of cellular engineering and offer several advantages compared to traditional gene circuits. However, engineering dynamic phenomena such as oscillations in protein-level circuits remains an outstanding challenge. Few examples of biological posttranslational oscillators are known, necessitating theoretical progress to determine realizable oscillators. We construct mathematical models for two posttranslational oscillators, using few components that interact only through reversible binding and phosphorylation/dephosphorylation reactions. Our designed oscillators rely on the self-assembly of two protein species into multimeric functional enzymes that respectively inhibit and enhance this self-assembly. We limit our analysis to within experimental constraints, finding (i) significant portions of the restricted parameter space yielding oscillations and (ii) that oscillation periods can be tuned by several orders of magnitude using recent advances in computational protein design. Our work paves the way for the rational design and realization of protein-based dynamic systems.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures



References
Publication types
LinkOut - more resources
Full Text Sources
