

Pharmacological Programming of Endosomal Signaling Activated by Small Molecule Ligands of the Follicle Stimulating Hormone Receptor
- PMID: 33329002
- PMCID: PMC7734412
- DOI: 10.3389/fphar.2020.593492
Pharmacological Programming of Endosomal Signaling Activated by Small Molecule Ligands of the Follicle Stimulating Hormone Receptor
Abstract
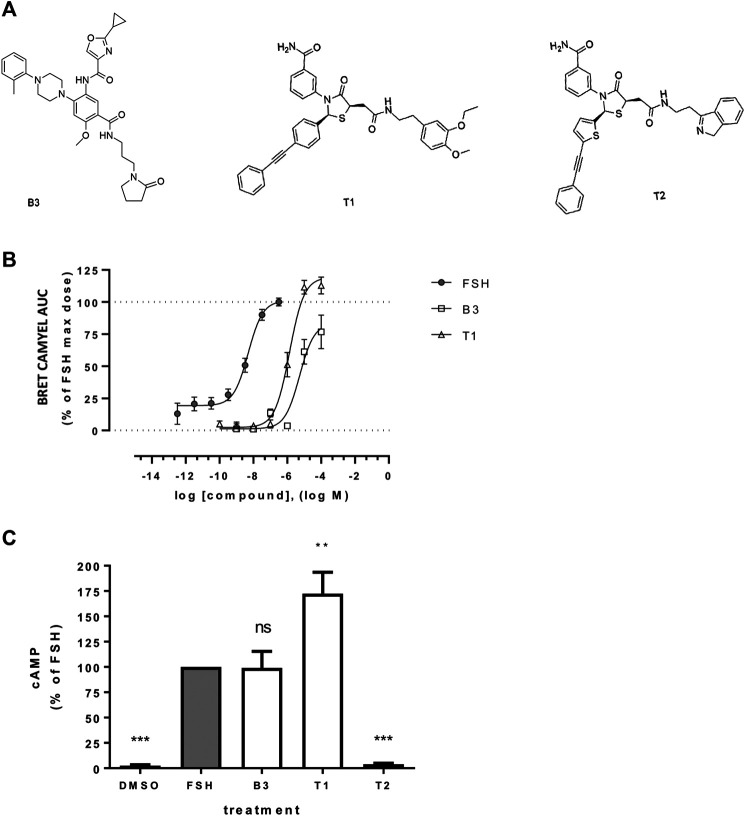
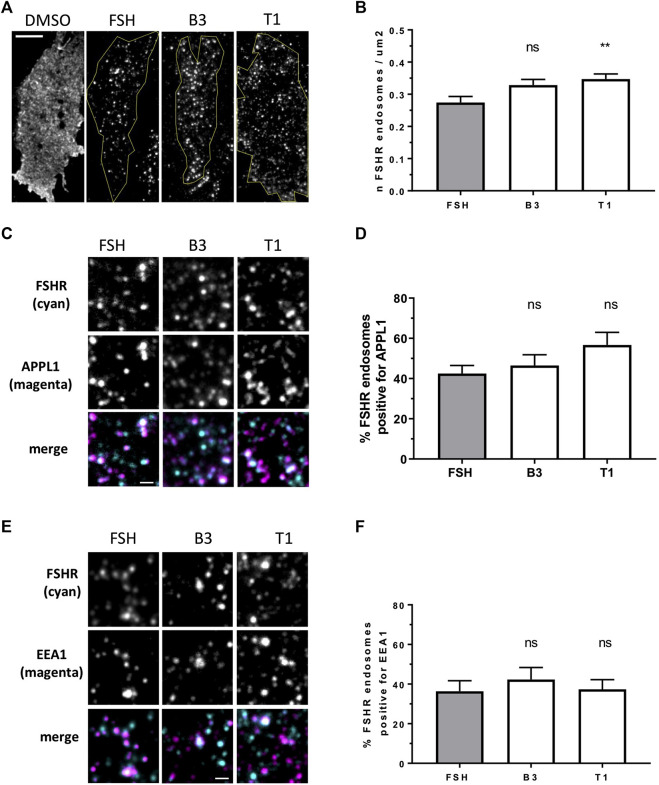
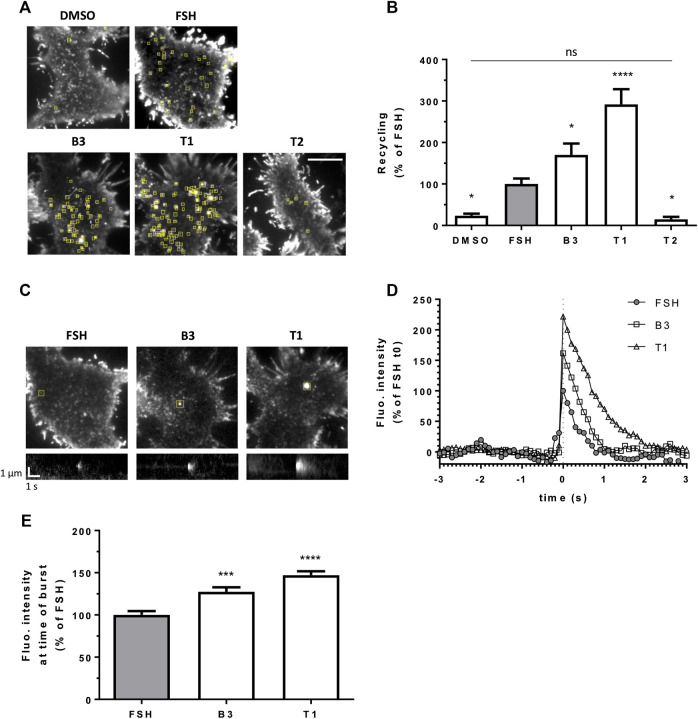
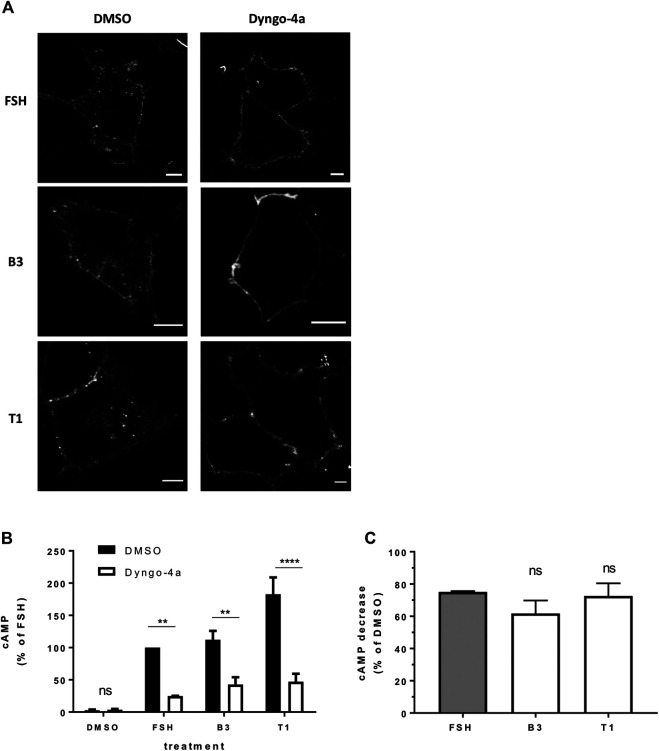
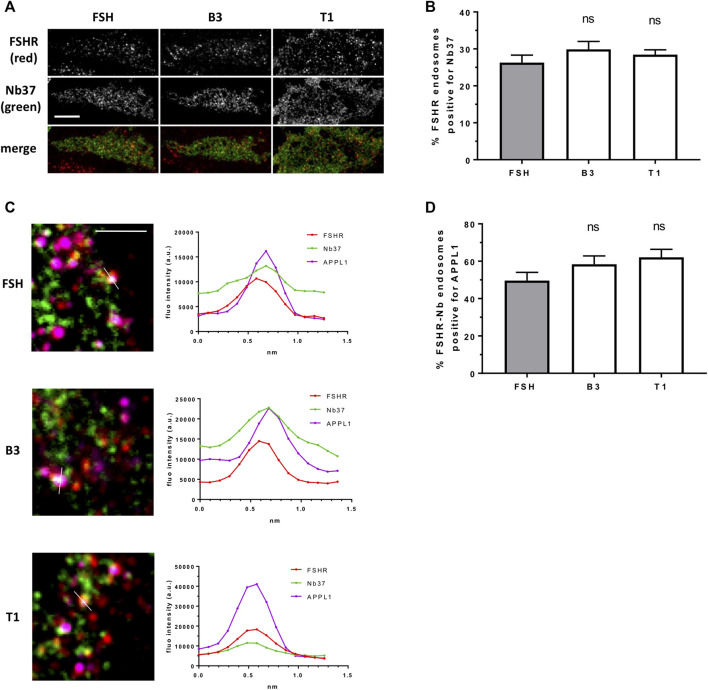
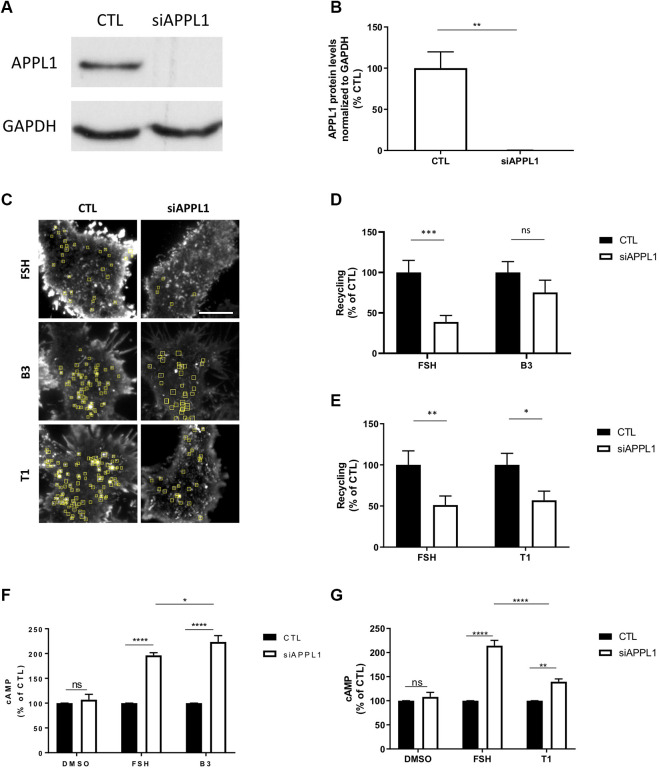
Follicle-stimulating hormone receptor (FSHR) is a G protein-coupled receptor (GPCR) with pivotal roles in reproduction. One key mechanism dictating the signal activity of GPCRs is membrane trafficking. After binding its hormone FSH, FSHR undergoes internalization to very early endosomes (VEEs) for its acute signaling and sorting to a rapid recycling pathway. The VEE is a heterogeneous compartment containing the Adaptor Protein Phosphotyrosine Interacting with Pleckstrin homology Domain and Leucine Zipper 1 (APPL1) with distinct functions in regulating endosomal Gαs/cAMP signaling and rapid recycling. Low molecular weight (LMW) allosteric FSHR ligands were developed for use in assisted reproductive technology yet could also provide novel pharmacological tools to study FSHR. Given the critical nature of receptor internalization and endosomal signaling for FSHR activity, we assessed whether these compounds exhibit differential abilities to alter receptor endosomal trafficking and signaling within the VEE. Two chemically distinct LMW agonists (benzamide, termed B3 and thiazolidinone, termed T1) were employed. T1 was able to induce a greater level of cAMP than FSH and B3. As cAMP signaling drives gonadotrophin hormone receptor recycling, rapid exocytic events were evaluated at single event resolution. Strikingly, T1 was able to induce a 3-fold increase in recycling events compared to FSH and two-fold more compared to B3. As T1-induced internalization was only marginally greater, the dramatic increase in recycling and cAMP signaling may be due to additional mechanisms. All compounds exhibited a similar requirement for receptor internalization to increase cAMP and proportion of FSHR endosomes with active Gαs, suggesting regulation of cAMP signaling induced by T1 may be altered. APPL1 plays a central role for GPCRs targeted to the VEE, and indeed, loss of APPL1 inhibited FSH-induced recycling and increased endosomal cAMP signaling. While T1-induced FSHR recycling was APPL1-dependent, its elevated cAMP signaling was only partially increased following APPL1 knockdown. Unexpectedly, B3 altered the dependence of FSHR to APPL1 in an opposing manner, whereby its endosomal signaling was negatively regulated by APPL1, while B3-induced FSHR recycling was APPL1-independent. Overall, FSHR allosteric compounds have the potential to re-program FSHR activity via altering engagement with VEE machinery and also suggests that these two distinct functions of APPL1 can potentially be selected pharmacologically.
Keywords: APPL1; G protein coupled receptor; allosteric ligand; endosome; follicle-stimulating hormone receptor.
Copyright © 2020 Sposini, De Pascali, Richardson, Sayers, Perrais, Yu, Palmer, Nataraja, Reiter and Hanyaloglu.
Conflict of interest statement
HY, SP, and SN were employees of TocopheRx when reagents were shared. Author SN is employed by the company Mitobridge Inc. Author HY is employed by the company Canwell Pharma. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials