

IGLR-2, a Leucine-Rich Repeat Domain Containing Protein, Is Required for the Host Defense in Caenorhabditis elegans
- PMID: 33329523
- PMCID: PMC7734252
- DOI: 10.3389/fimmu.2020.561337
IGLR-2, a Leucine-Rich Repeat Domain Containing Protein, Is Required for the Host Defense in Caenorhabditis elegans
Abstract
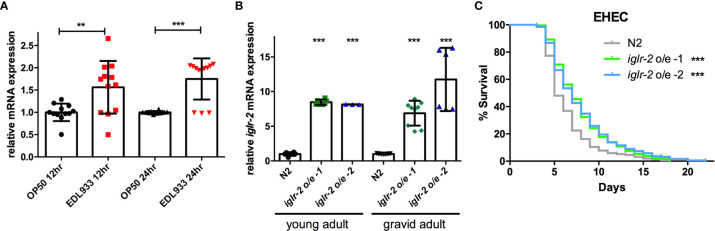
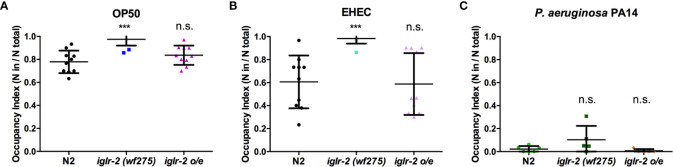
Enterohemorrhagic Escherichia coli (EHEC), a human pathogen, also infects Caenorhabditis elegans. We demonstrated previously that C. elegans activates the p38 MAPK innate immune pathway to defend against EHEC infection. However, whether a C. elegans pattern recognition receptor (PRR) exists to regulate the immune pathway remains unknown. PRRs identified in other metazoans contain several conserved domains, including the leucine-rich repeat (LRR). By screening a focused RNAi library, we identified the IGLR-2, a transmembrane protein containing the LRR domain, as a potential immune regulator in C. elegans. Our data showed that iglr-2 regulates the host susceptibility to EHEC infection. Moreover, iglr-2 is required for pathogen avoidance to EHEC. The iglr-2 overexpressed strain, which was more resistant to EHEC originally, showed hypersusceptibility to EHEC upon knockdown of the p38 MAPK pathway. Together, our data suggested that iglr-2 plays an important role in C. elegans to defend EHEC by regulating pathogen-avoidance behavior and the p38 MAPK pathway.
Keywords: Caenorhabditis elegans; enterohemorrhagic Escherichia coli; iglr-2; innate immunity; p38 MAPK pathway.
Copyright © 2020 Kuo, Hsu, Wang, Liou, Lim, Chen and Chen.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
The conserved regulator of autophagy and innate immunity hlh-30/TFEB mediates tolerance of enterohemorrhagic Escherichia coli in Caenorhabditis elegans.Genetics. 2021 Mar 3;217(1):1-17. doi: 10.1093/genetics/iyaa052. Genetics. 2021. PMID: 33683370 Free PMC article.
-
Enterohaemorrhagic Escherichia coli O157:H7 Shiga-like toxin 1 is required for full pathogenicity and activation of the p38 mitogen-activated protein kinase pathway in Caenorhabditis elegans.Cell Microbiol. 2013 Jan;15(1):82-97. doi: 10.1111/cmi.12030. Epub 2012 Oct 9. Cell Microbiol. 2013. PMID: 22985085
-
Mutation of the Enterohemorrhagic Escherichia coli Core LPS Biosynthesis Enzyme RfaD Confers Hypersusceptibility to Host Intestinal Innate Immunity In vivo.Front Cell Infect Microbiol. 2016 Aug 12;6:82. doi: 10.3389/fcimb.2016.00082. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27570746 Free PMC article.
-
Evolutionary perspectives on innate immunity from the study of Caenorhabditis elegans.Curr Opin Immunol. 2005 Feb;17(1):4-10. doi: 10.1016/j.coi.2004.11.007. Curr Opin Immunol. 2005. PMID: 15653303 Review.
-
[Progress in the mechanisms of neural modulation of innate immunity in Caenorhabditis elegans].Yi Chuan. 2018 Dec 20;40(12):1066-1074. doi: 10.16288/j.yczz.18-133. Yi Chuan. 2018. PMID: 30559096 Review. Chinese.
Cited by
-
Detection of Pseudomonas aeruginosa Serogroup G Using Real-Time PCR for Novel Target Genes Identified Through Comparative Genomics.Front Microbiol. 2022 Jun 24;13:928154. doi: 10.3389/fmicb.2022.928154. eCollection 2022. Front Microbiol. 2022. PMID: 35814691 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials