

Lymph nodes are innervated by a unique population of sensory neurons with immunomodulatory potential
- PMID: 33333021
- PMCID: PMC9612289
- DOI: 10.1016/j.cell.2020.11.028
Lymph nodes are innervated by a unique population of sensory neurons with immunomodulatory potential
Abstract
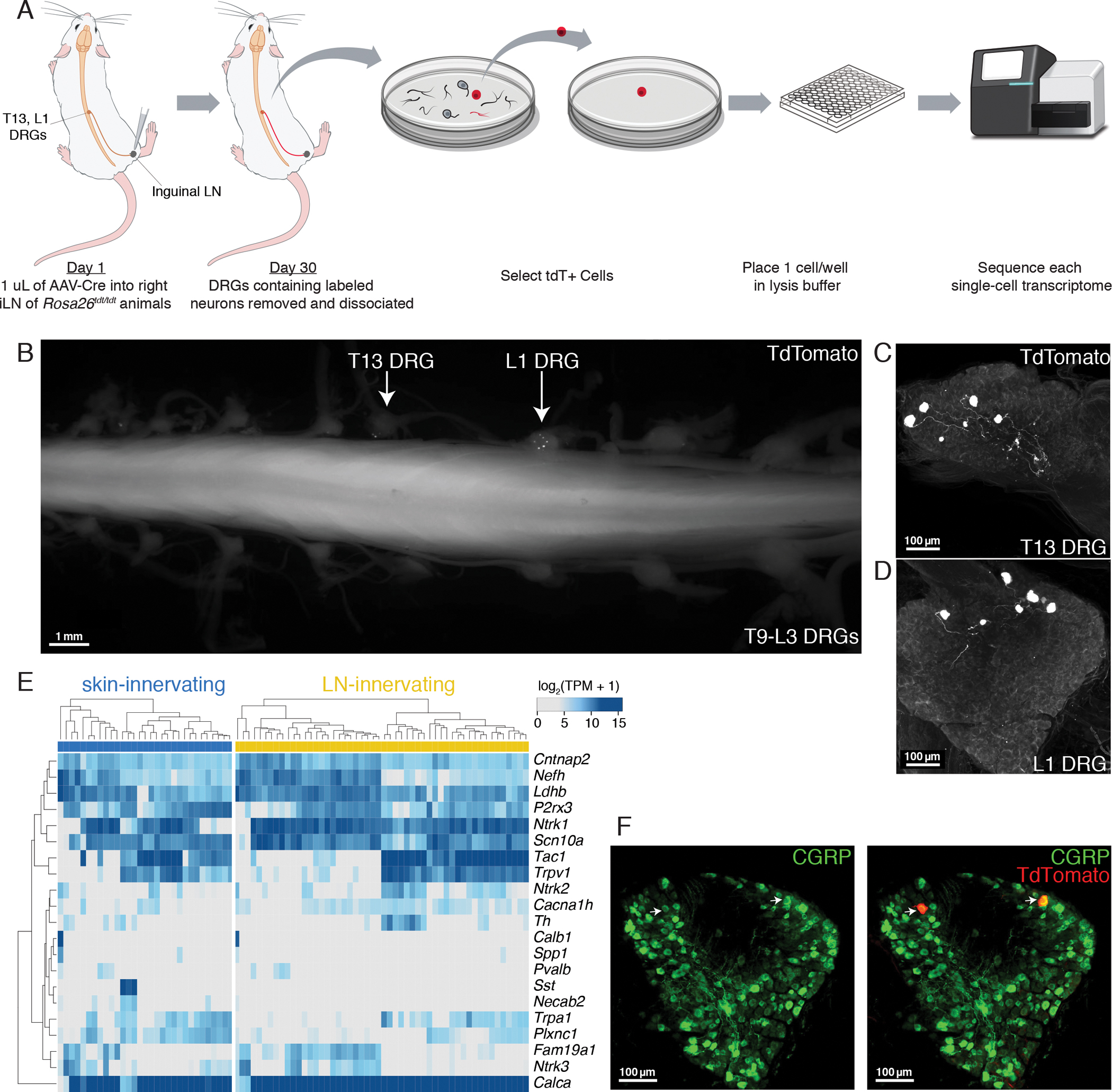
Barrier tissue immune responses are regulated in part by nociceptors. Nociceptor ablation alters local immune responses at peripheral sites and within draining lymph nodes (LNs). The mechanisms and significance of nociceptor-dependent modulation of LN function are unknown. Using high-resolution imaging, viral tracing, single-cell transcriptomics, and optogenetics, we identified and functionally tested a sensory neuro-immune circuit that is responsive to lymph-borne inflammatory signals. Transcriptomics profiling revealed that multiple sensory neuron subsets, predominantly peptidergic nociceptors, innervate LNs, distinct from those innervating surrounding skin. To uncover LN-resident cells that may interact with LN-innervating sensory neurons, we generated a LN single-cell transcriptomics atlas and nominated nociceptor target populations and interaction modalities. Optogenetic stimulation of LN-innervating sensory fibers triggered rapid transcriptional changes in the predicted interacting cell types, particularly endothelium, stromal cells, and innate leukocytes. Thus, a unique population of sensory neurons monitors peripheral LNs and may locally regulate gene expression.
Keywords: lymph node; neuro-immunology; nociceptor; optogenetics; sensory neuron; single-cell RNA-seq.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.K.S. has received compensation for consulting and SAB membership from Honeycomb Biotechnologies, Cellarity, Repertoire Immune Medicines, Ochre Bio, and Dahlia Biosciences. U.H.v.A. has received compensation for consulting and SAB membership from Beam, Cogen Therapeutics, Cygnal, Moderna, Monopteros, Morphic, Rubius, Selecta Biosciences, SQZ, and Synlogic. S.H., C.G.K.Z., A.K.S., and U.H.v.A. are co-inventors on a provisional patent application filed by the Broad Institute (U.S. patent no. 62/916,184) relating to the results described in this manuscript.
Figures






Comment in
-
Sensory Nerves Regulate Transcriptional Dynamics of Lymph Node Cells.Trends Immunol. 2021 Mar;42(3):180-182. doi: 10.1016/j.it.2021.01.008. Epub 2021 Feb 6. Trends Immunol. 2021. PMID: 33563563
References
-
- Abrahamsen B, Zhao J, Asante CO, Cendan CM, Marsh S, Martinez-Barbera JP, Nassar MA, Dickenson AH, and Wood JN (2008). The cell and molecular basis of mechanical, cold, and inflammatory pain. Science 321, 702–705. - PubMed
-
- Akira S, Uematsu S, and Takeuchi O (2006). Pathogen recognition and innate immunity. Cell 124, 783–801. - PubMed
-
- Andoh T, Nishikawa Y, Yamaguchi-Miyamoto T, Nojima H, Narumiya S, and Kuraishi Y (2007). Thromboxane A2 induces itch-associated responses through TP receptors in the skin in mice. J Invest Dermatol 127, 2042–2047. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
