

Large Multidomain Protein NMR: HIV-1 Reverse Transcriptase Precursor in Solution
- PMID: 33333923
- PMCID: PMC7765405
- DOI: 10.3390/ijms21249545
Large Multidomain Protein NMR: HIV-1 Reverse Transcriptase Precursor in Solution
Abstract
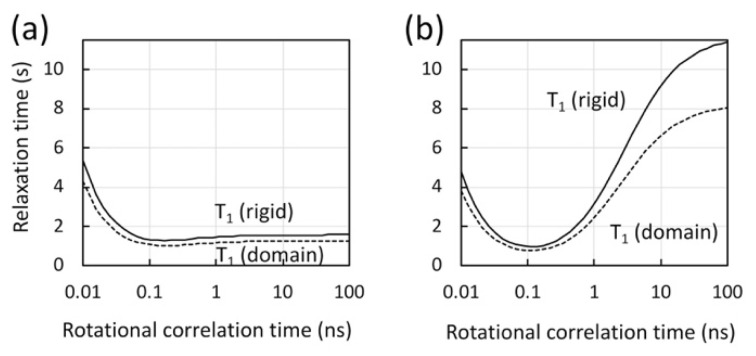
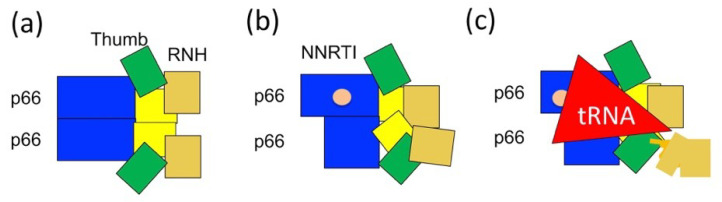
NMR studies of large proteins, over 100 kDa, in solution are technically challenging and, therefore, of considerable interest in the biophysics field. The challenge arises because the molecular tumbling of a protein in solution considerably slows as molecular mass increases, reducing the ability to detect resonances. In fact, the typical 1H-13C or 1H-15N correlation spectrum of a large protein, using a 13C- or 15N-uniformly labeled protein, shows severe line-broadening and signal overlap. Selective isotope labeling of methyl groups is a useful strategy to reduce these issues, however, the reduction in the number of signals that goes hand-in-hand with such a strategy is, in turn, disadvantageous for characterizing the overall features of the protein. When domain motion exists in large proteins, the domain motion differently affects backbone amide signals and methyl groups. Thus, the use of multiple NMR probes, such as 1H, 19F, 13C, and 15N, is ideal to gain overall structural or dynamical information for large proteins. We discuss the utility of observing different NMR nuclei when characterizing a large protein, namely, the 66 kDa multi-domain HIV-1 reverse transcriptase that forms a homodimer in solution. Importantly, we present a biophysical approach, complemented by biochemical assays, to understand not only the homodimer, p66/p66, but also the conformational changes that contribute to its maturation to a heterodimer, p66/p51, upon HIV-1 protease cleavage.
Keywords: HIV; NMR; isotope labeling; protein; reverse transcriptase; ribonuclease H.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Spatial domain organization in the HIV-1 reverse transcriptase p66 homodimer precursor probed by double electron-electron resonance EPR.Proc Natl Acad Sci U S A. 2019 Sep 3;116(36):17809-17816. doi: 10.1073/pnas.1911086116. Epub 2019 Aug 5. Proc Natl Acad Sci U S A. 2019. PMID: 31383767 Free PMC article.
-
Conformational Changes in HIV-1 Reverse Transcriptase that Facilitate Its Maturation.Structure. 2019 Oct 1;27(10):1581-1593.e3. doi: 10.1016/j.str.2019.08.004. Epub 2019 Aug 27. Structure. 2019. PMID: 31471129 Free PMC article.
-
The HIV-1 p66 homodimeric RT exhibits different conformations in the binding-competent and -incompetent NNRTI site.Proteins. 2017 Dec;85(12):2191-2197. doi: 10.1002/prot.25383. Epub 2017 Sep 26. Proteins. 2017. PMID: 28905420 Free PMC article.
-
HIV-1 Reverse Transcriptase: A Metamorphic Protein with Three Stable States.Structure. 2019 Mar 5;27(3):420-426. doi: 10.1016/j.str.2018.11.011. Epub 2019 Jan 10. Structure. 2019. PMID: 30639227 Free PMC article. Review.
-
Conformational changes in HIV-1 reverse transcriptase induced by nonnucleoside reverse transcriptase inhibitor binding.Curr HIV Res. 2004 Oct;2(4):323-32. doi: 10.2174/1570162043351093. Curr HIV Res. 2004. PMID: 15544453 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
