

JNK2, a Newly-Identified SERCA2 Enhancer, Augments an Arrhythmic [Ca2+]SR Leak-Load Relationship
- PMID: 33334123
- PMCID: PMC7897290
- DOI: 10.1161/CIRCRESAHA.120.318409
JNK2, a Newly-Identified SERCA2 Enhancer, Augments an Arrhythmic [Ca2+]SR Leak-Load Relationship
Abstract
Rationale: We recently discovered pivotal contributions of stress kinase JNK2 (c-Jun N-terminal kinase isoform 2) in increased risk of atrial fibrillation through enhanced diastolic sarcoplasmic reticulum (SR) calcium (Ca2+) leak via RyR2 (ryanodine receptor isoform 2). However, the role of JNK2 in the function of the SERCA2 (SR Ca2+-ATPase), essential in maintaining SR Ca2+ content cycling during each heartbeat, is completely unknown.
Objective: To test the hypothesis that JNK2 increases SERCA2 activity SR Ca2+ content and exacerbates an arrhythmic SR Ca2+ content leak-load relationship.
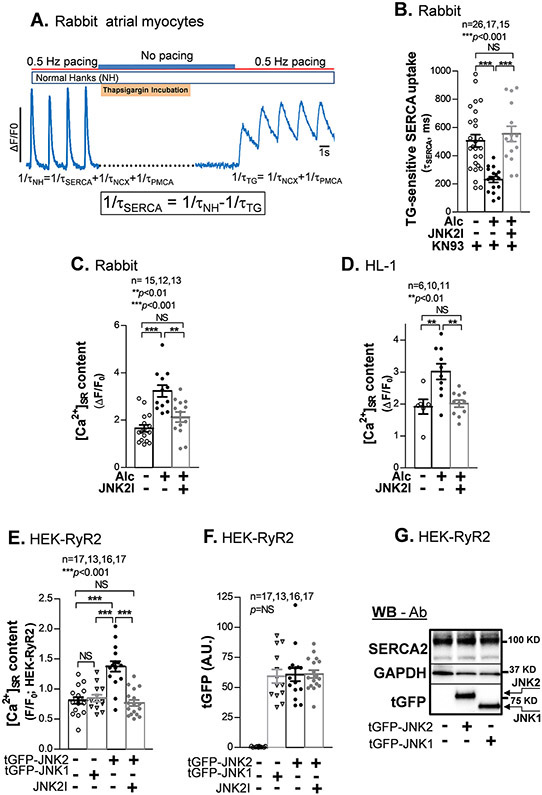
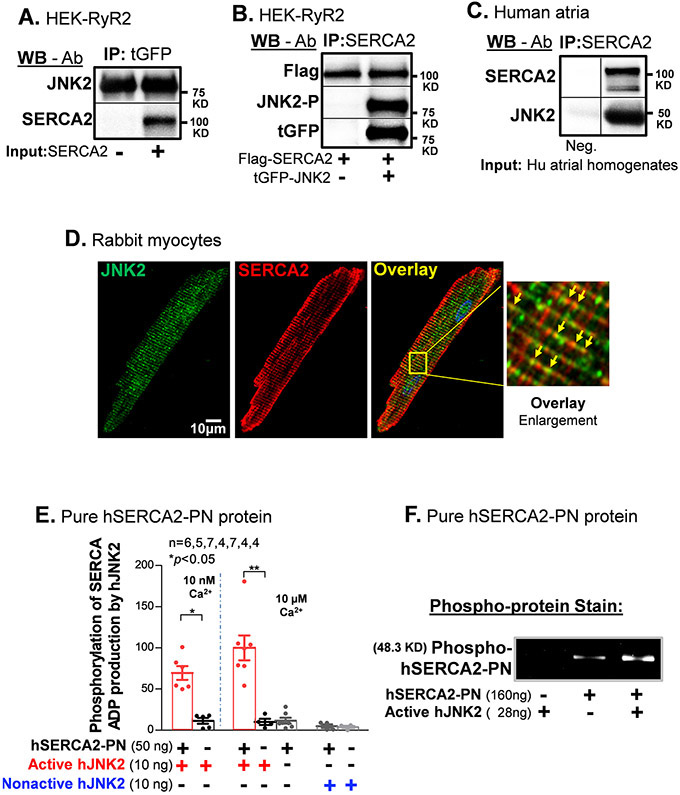
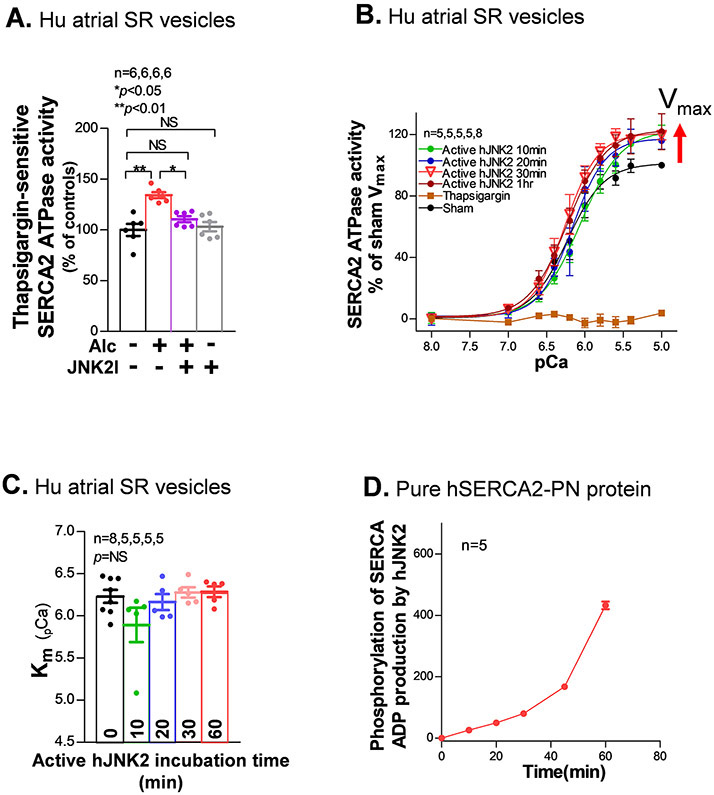
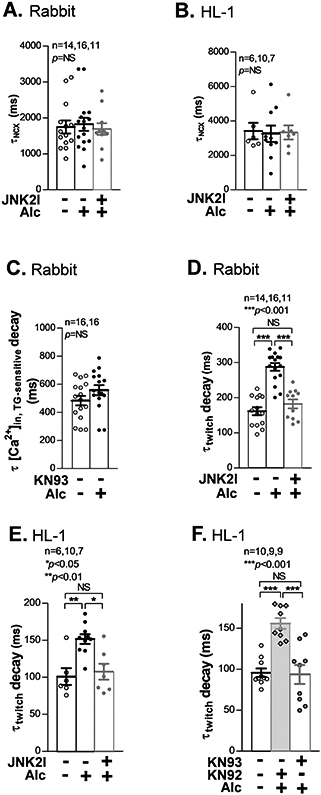
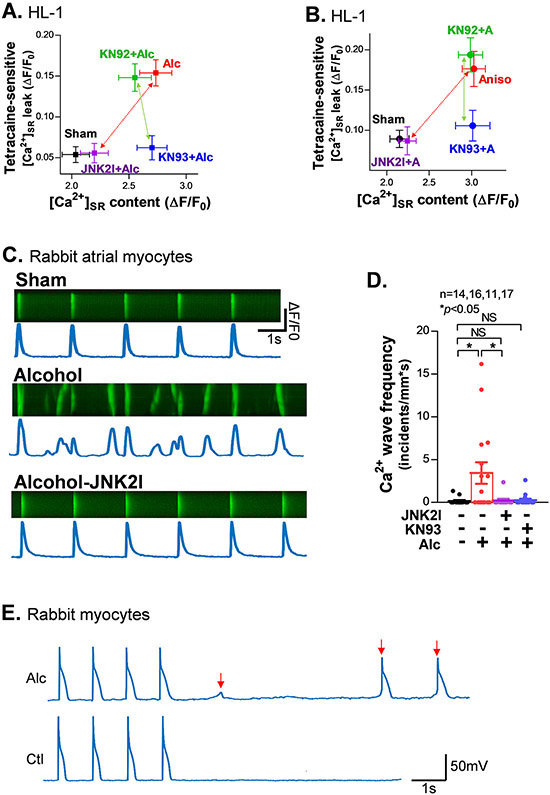
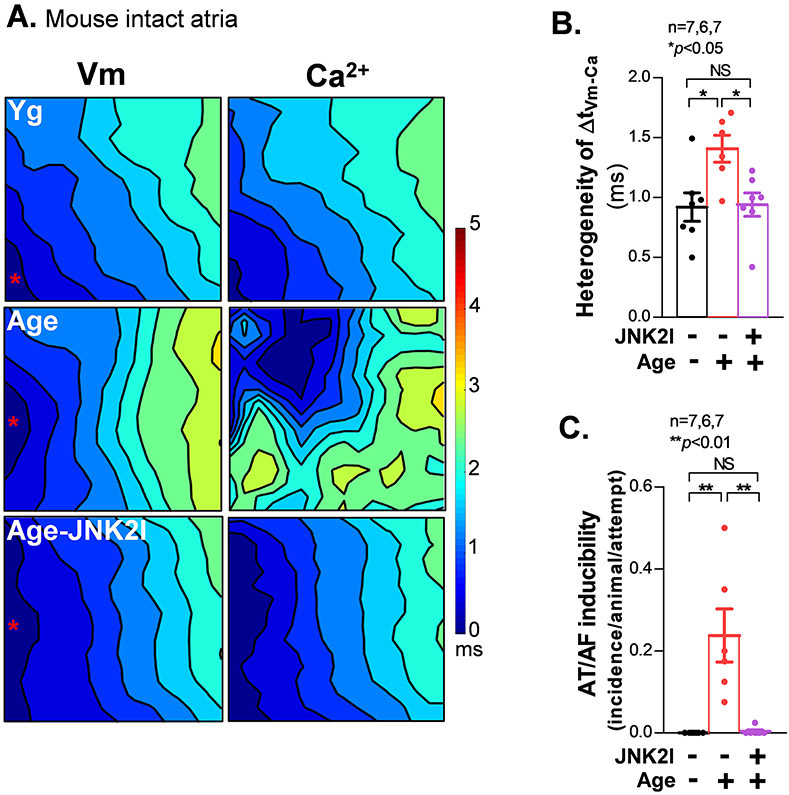
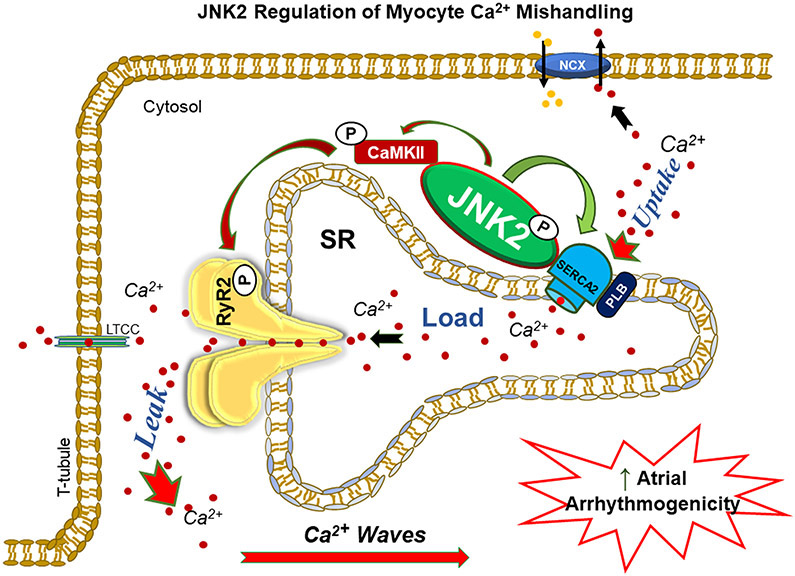
Methods and results: We used confocal Ca2+ imaging in myocytes and HEK-RyR2 (ryanodine receptor isoform 2-expressing human embryonic kidney 293 cells) cells, biochemistry, dual Ca2+/voltage optical mapping in intact hearts from alcohol-exposed or aged mice (where JNK2 is activated). We found that JNK2, but not JNK1 (c-Jun N-terminal kinase isoform 1), increased SERCA2 uptake and consequently elevated SR Ca2+ content load. JNK2 also associates with and phosphorylates SERCA2 proteins. JNK2 causally enhances SERCA2-ATPase activity via increased maximal rate, without altering Ca2+ affinity. Unlike the CaMKII (Ca2+/calmodulin-dependent kinase II)-dependent JNK2 action in SR Ca2+ leak, JNK2-driven SERCA2 function was CaMKII independent (not prevented by CaMKII inhibition). With CaMKII blocked, the JNK2-driven SR Ca2+ loading alone did not significantly raise leak. However, with JNK2-CaMKII-driven SR Ca2+ leak present, the JNK2-enhanced SR Ca2+ uptake limited leak-induced reduction in SR Ca2+, normalizing Ca2+ transient amplitude, but at a higher arrhythmogenic SR Ca2+ leak. JNK2-specific inhibition completely normalized SR Ca2+ handling, attenuated arrhythmic Ca2+ activities, and alleviated atrial fibrillation susceptibility in aged and alcohol-exposed myocytes and intact hearts.
Conclusions: We have identified a novel JNK2-induced activation of SERCA2. The dual action of JNK2 in CaMKII-dependent arrhythmic SR Ca2+ leak and a CaMKII-independent uptake exacerbates atrial arrhythmogenicity, while helping to maintain normal levels of Ca2+ transients and heart function. JNK2 modulation may be a novel therapeutic target for atrial fibrillation prevention and treatment.
Keywords: JNK mitogen-activated protein kinases; animals; atrial fibrillation; phosphorylation; sarcoplasmic reticulum.
Figures

Comment in
-
Stress Seats a New Guest at the Table of PLN/SERCA and Their Partners.Circ Res. 2021 Feb 19;128(4):471-473. doi: 10.1161/CIRCRESAHA.121.318742. Epub 2021 Feb 18. Circ Res. 2021. PMID: 33600231 Free PMC article. No abstract available.
References
-
- Go AS, Hylek EM, Phillips KA, Chang Y, Henault LE, Selby JV and Singer DE. Prevalence of diagnosed atrial fibrillation in adults: national implications for rhythm management and stroke prevention: the AnTicoagulation and Risk Factors in Atrial Fibrillation (ATRIA) Study. JAMA. 2001;285:2370–5. - PubMed
-
- Benjamin EJ, Levy D, Vaziri SM, D'Agostino RB, Belanger AJ and Wolf PA. Independent risk factors for atrial fibrillation in a population-based cohort. The Framingham Heart Study. JAMA. 1994;271:840–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
