

A Critical Role for Fas-Mediated Off-Target Tumor Killing in T-cell Immunotherapy
- PMID: 33334730
- PMCID: PMC7933082
- DOI: 10.1158/2159-8290.CD-20-0756
A Critical Role for Fas-Mediated Off-Target Tumor Killing in T-cell Immunotherapy
Abstract
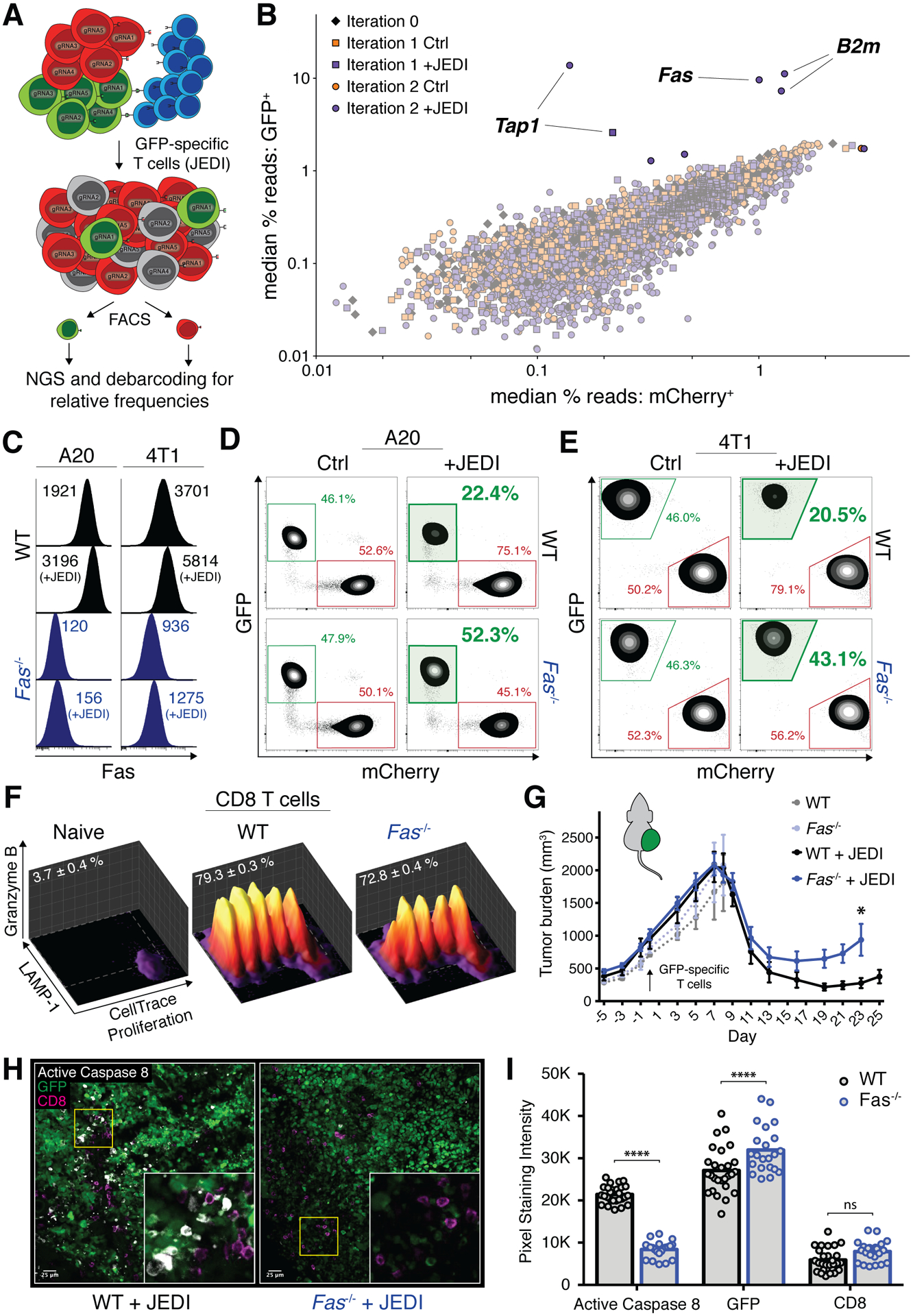
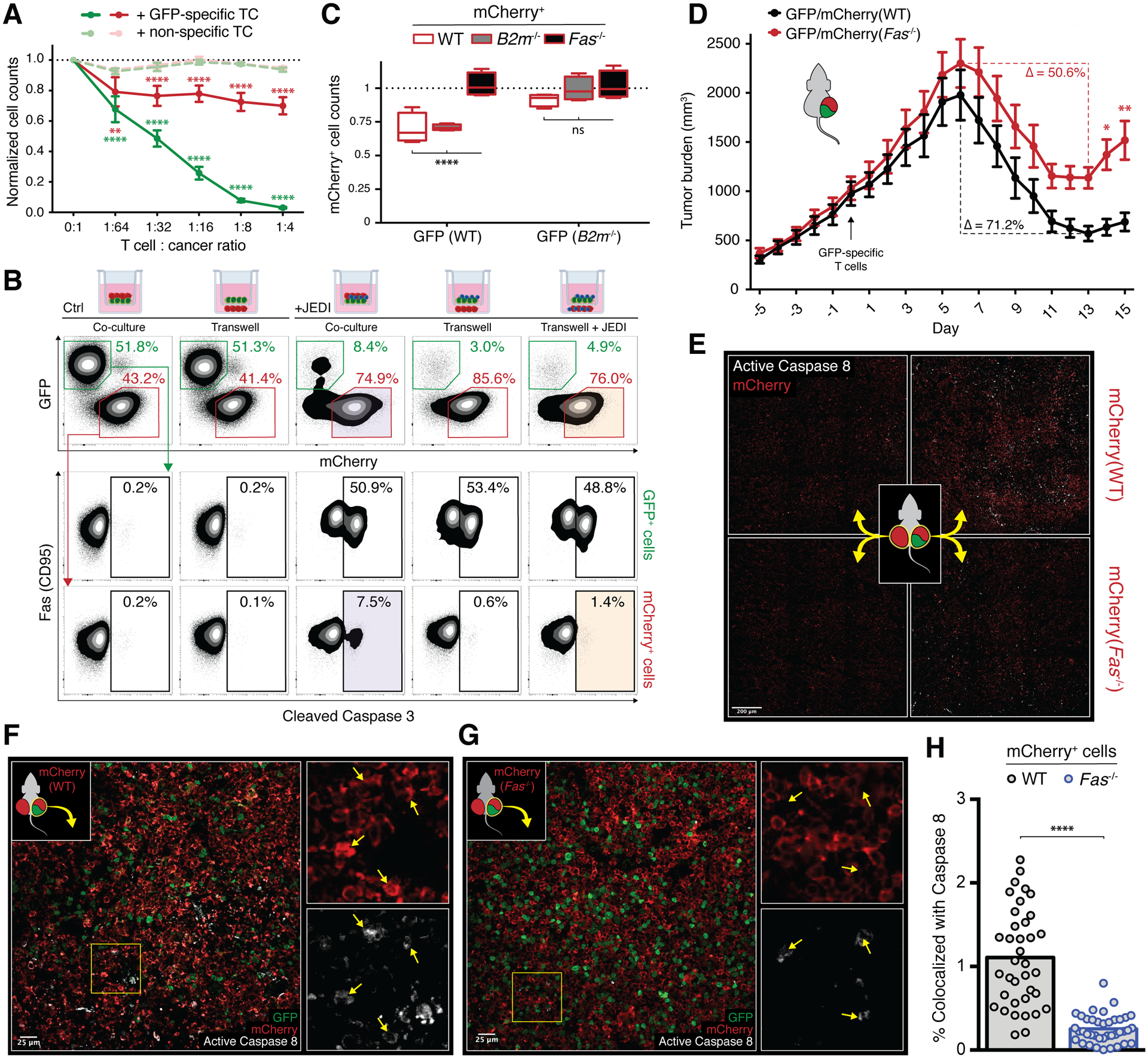
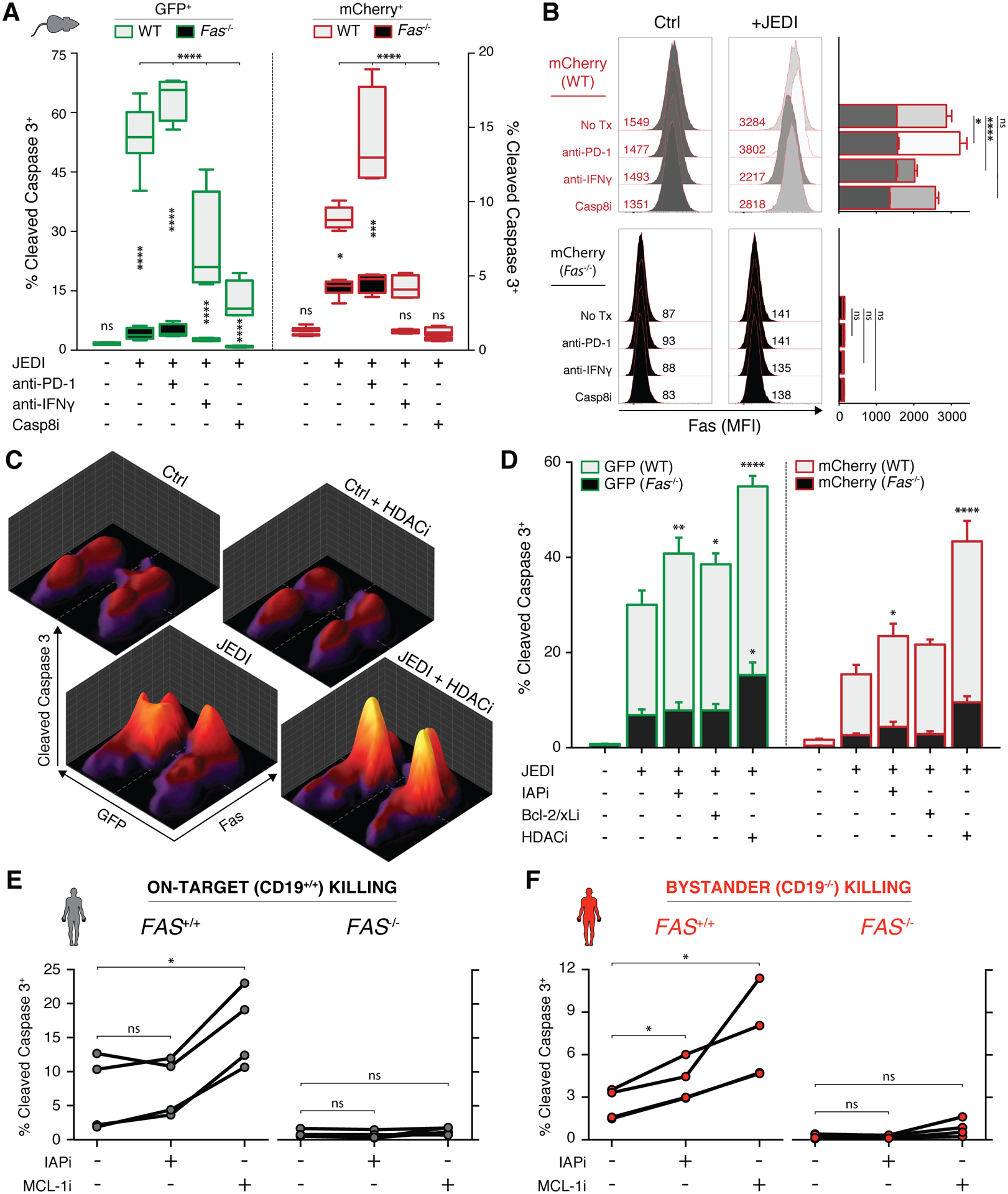
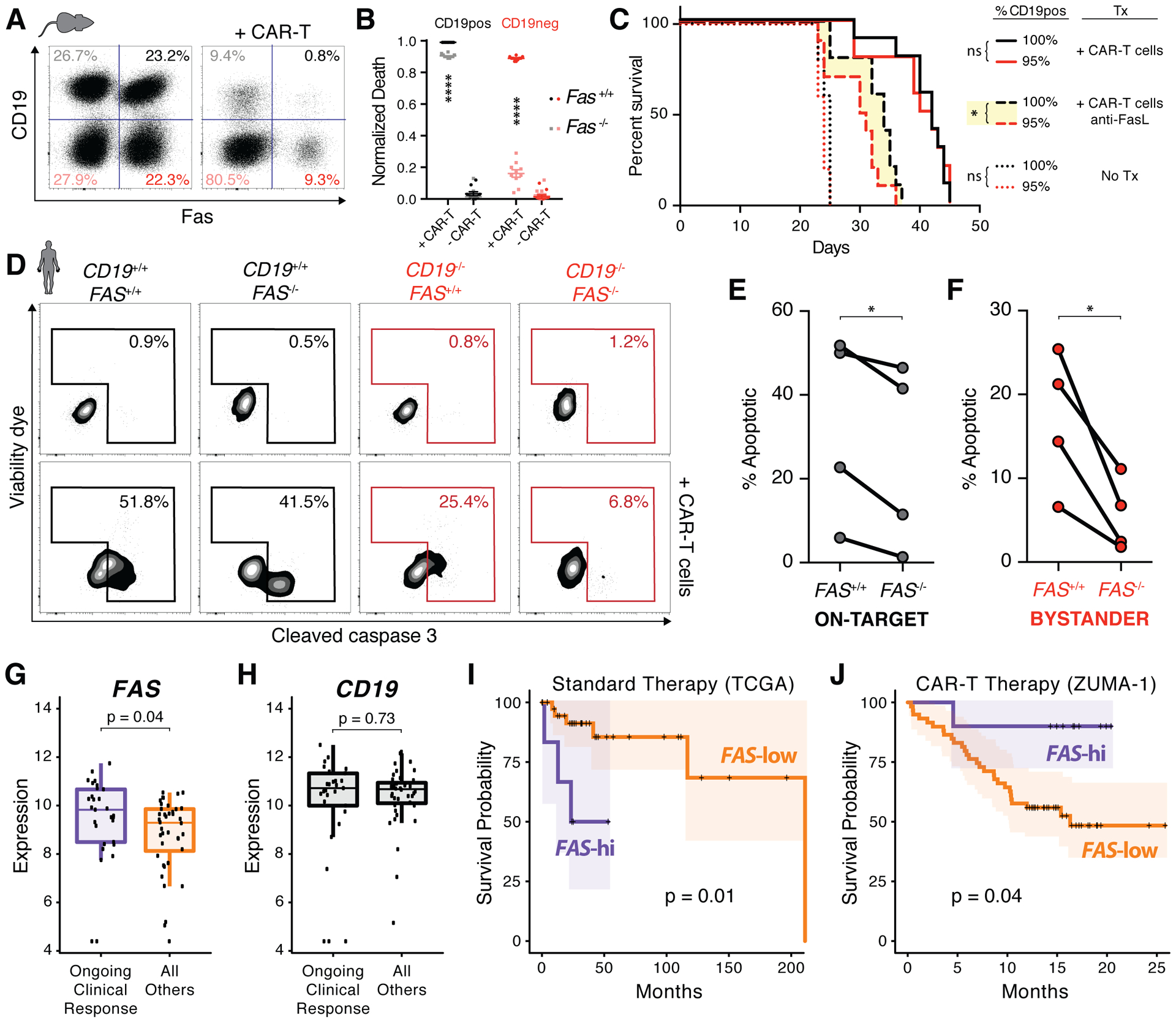
T cell-based therapies have induced cancer remissions, though most tumors ultimately progress, reflecting inherent or acquired resistance including antigen escape. Better understanding of how T cells eliminate tumors will help decipher resistance mechanisms. We used a CRISPR/Cas9 screen and identified a necessary role for Fas-FasL in antigen-specific T-cell killing. We also found that Fas-FasL mediated off-target "bystander" killing of antigen-negative tumor cells. This localized bystander cytotoxicity enhanced clearance of antigen-heterogeneous tumors in vivo, a finding that has not been shown previously. Fas-mediated on-target and bystander killing was reproduced in chimeric antigen receptor (CAR-T) and bispecific antibody T-cell models and was augmented by inhibiting regulators of Fas signaling. Tumoral FAS expression alone predicted survival of CAR-T-treated patients in a large clinical trial (NCT02348216). These data suggest strategies to prevent immune escape by targeting both the antigen expression of most tumor cells and the geography of antigen-loss variants. SIGNIFICANCE: This study demonstrates the first report of in vivo Fas-dependent bystander killing of antigen-negative tumors by T cells, a phenomenon that may be contributing to the high response rates of antigen-directed immunotherapies despite tumoral heterogeneity. Small molecules that target the Fas pathway may potentiate this mechanism to prevent cancer relapse.This article is highlighted in the In This Issue feature, p. 521.
©2020 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Hammerich L, Marron TU, Upadhyay R, Svensson-Arvelund J, Dhainaut M, Hussein S, et al. Systemic clinical tumor regressions and potentiation of PD1 blockade with in situ vaccination. Nat Med 2019;25:814–24. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
