

Phagocytic function of tumor-associated macrophages as a key determinant of tumor progression control: a review
- PMID: 33335026
- PMCID: PMC7747550
- DOI: 10.1136/jitc-2020-001408
Phagocytic function of tumor-associated macrophages as a key determinant of tumor progression control: a review
Abstract
Tumor-associated macrophage (TAM) phagocytic activity is emerging as a new mechanism to harness for cancer treatment. Currently, many approaches are investigated at the preclinical level and some modalities have now reached clinical trials, including the targeting of the phagocytosis inhibitor CD47. The rationale for increasing TAM phagocytic activity is to improve innate anticancer immunity, and to promote T-cell mediated adaptive immune responses. In this context, a clear understanding of the impact of TAM phagocytosis on both innate and adaptive immunity is critical. Indeed, uncertainties persist regarding the capacity of TAM to present tumor antigens to CD8 T cells by cross-presentation. This process is critical for an optimal cytotoxic T-cell immune response and can be mediated by dendritic cells but also potentially by macrophages. In addition, the engulfment of cancer cells affects TAM functionality, as apoptotic cell uptake (a process termed efferocytosis) promotes macrophage anti-inflammatory functions. Because of the abundance of TAM in most solid tumors and the common use of apoptosis inducers such as radiotherapy to treat patients with cancer, efferocytosis potentially affects the overall immune balance within the tumor microenvironment (TME). In this review, we will discuss how cancer cell phagocytosis by TAM impacts antitumor immunity. First, we will focus on the potential of the phagocytic activity of TAM per se to control tumor progression. Second, we will examine the potential of TAM to act as antigen presenting cells for tumor specific CD8 T cells, considering the different characteristics of this process in the tumor tissue and at the molecular level. Finally, we will see how phagocytosis and efferocytosis affect TAM functionality and how these mechanisms impact on antitumor immunity. A better understanding of these aspects will enable us to better predict and interpret the consequences of cancer therapies on the immune status of the TME. Future cancer treatment regimens can thereby be designed to not only impact directly on cancer cells, but also to favorably modulate TAM phagocytic activity to benefit from the potential of this central immune player to achieve more potent therapeutic efficacy.
Keywords: antigen presentation; immunity; innate; macrophages; phagocytosis; tumor microenvironment.
© Author(s) (or their employer(s)) 2020. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures
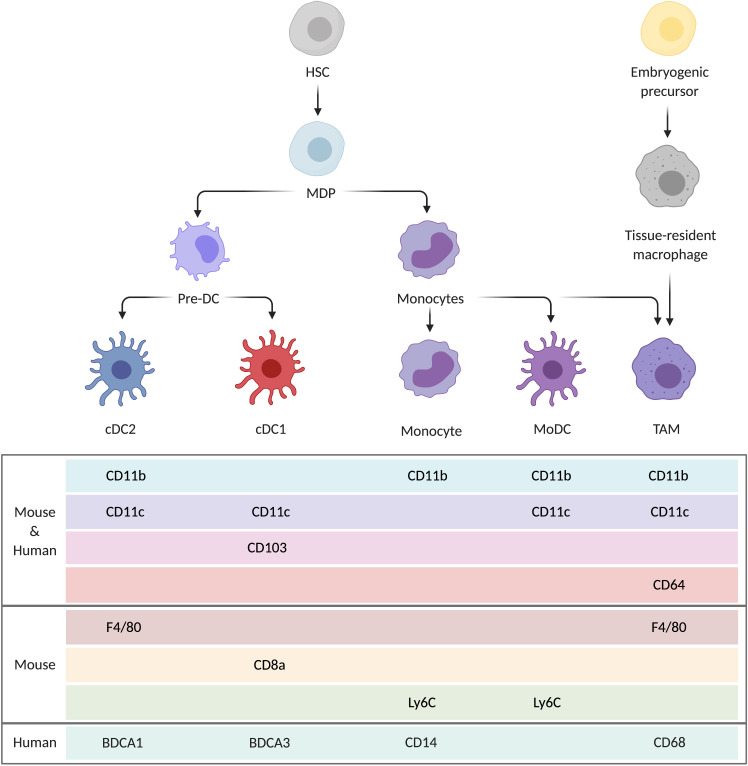
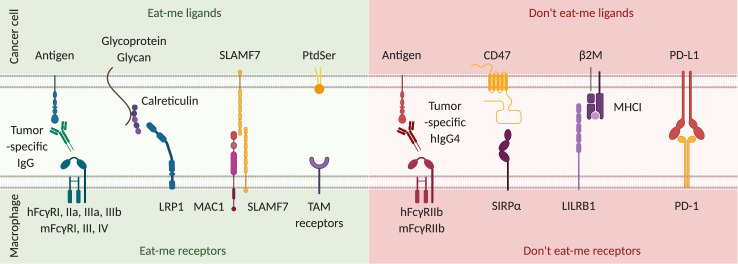
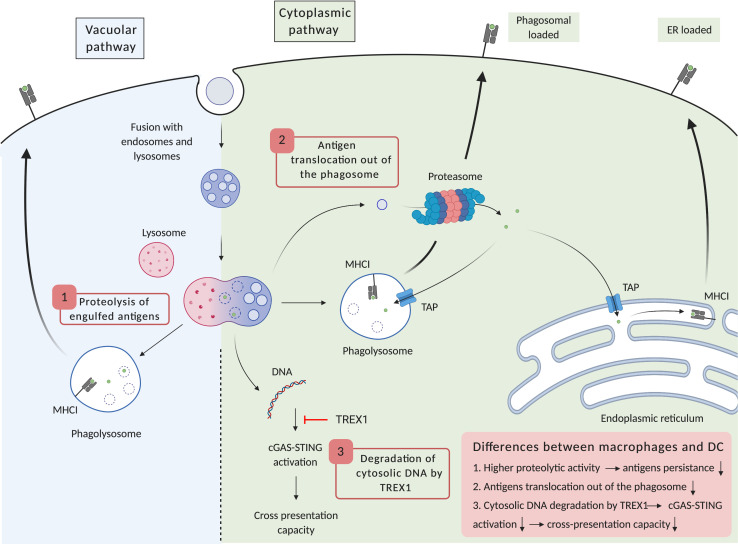
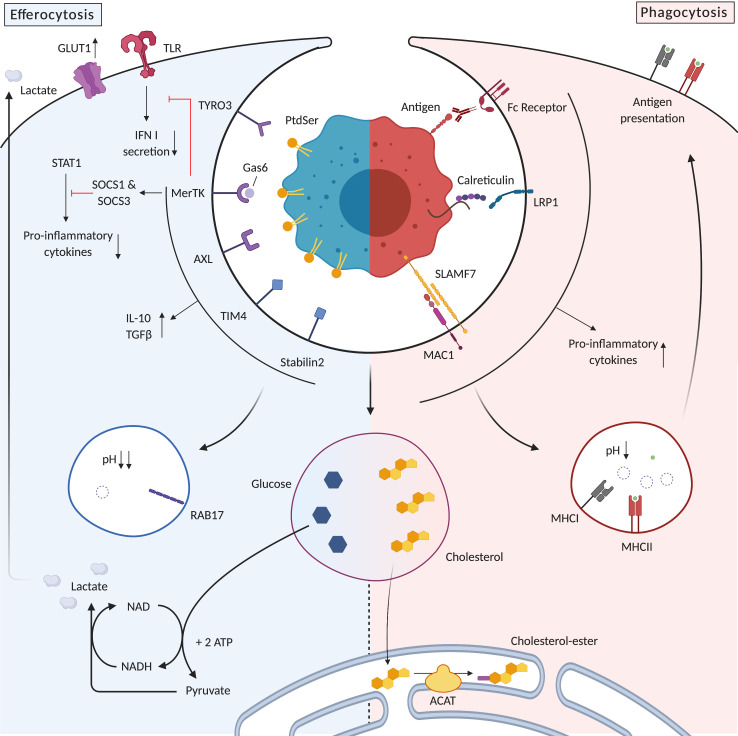
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials