

Cerebellar nuclei evolved by repeatedly duplicating a conserved cell-type set
- PMID: 33335034
- PMCID: PMC8510508
- DOI: 10.1126/science.abd5059
Cerebellar nuclei evolved by repeatedly duplicating a conserved cell-type set
Abstract
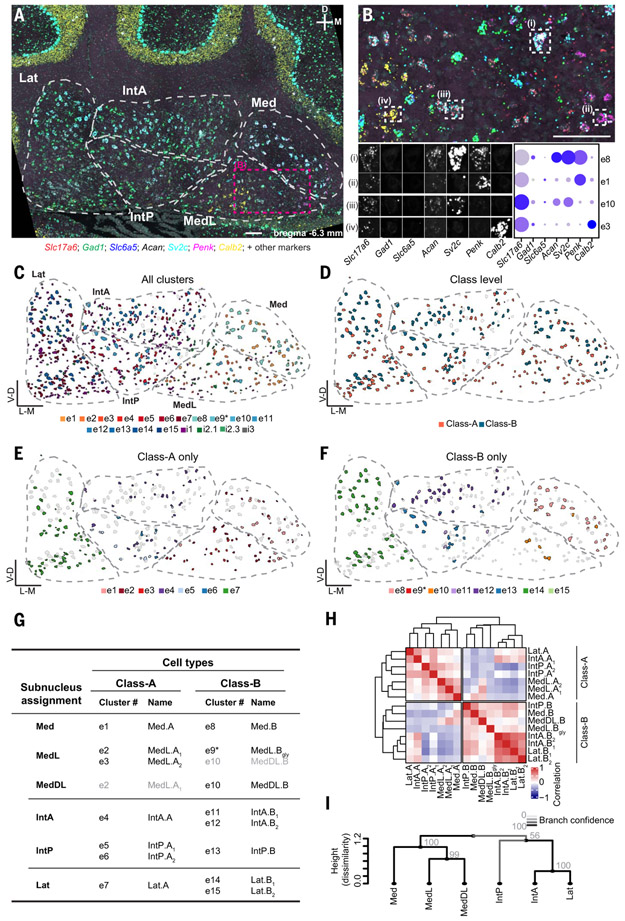
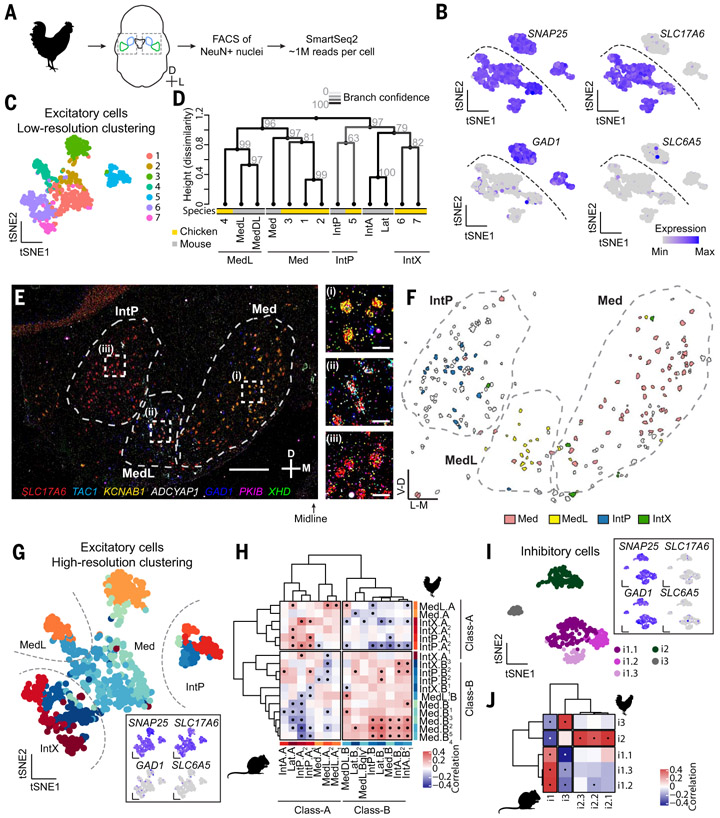
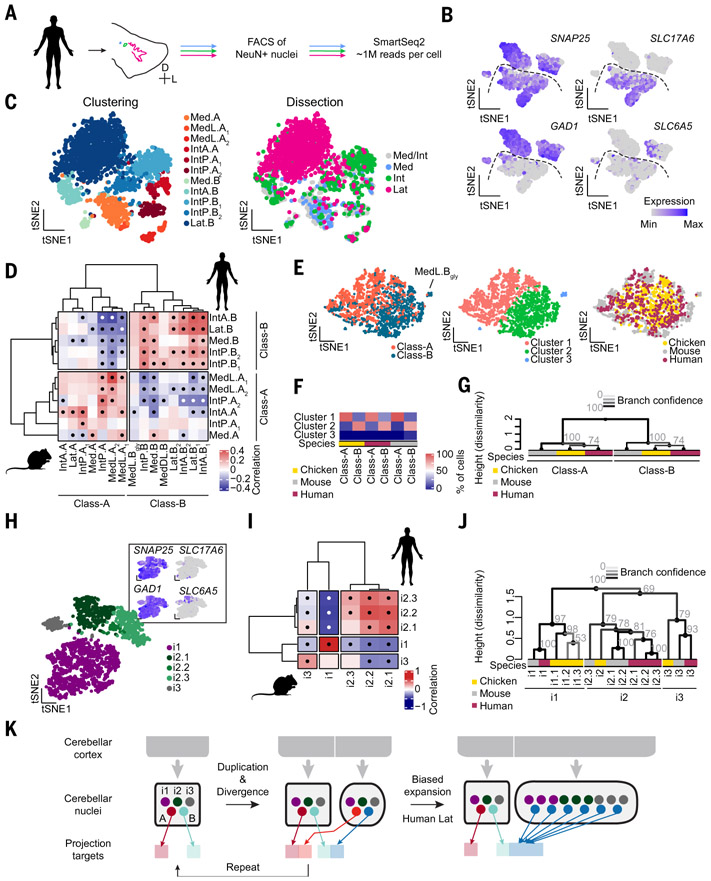
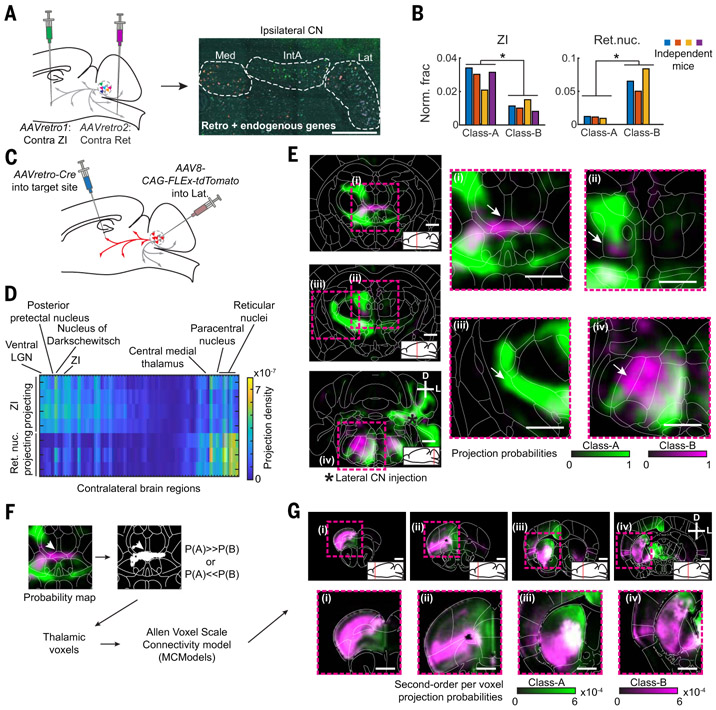
How have complex brains evolved from simple circuits? Here we investigated brain region evolution at cell-type resolution in the cerebellar nuclei, the output structures of the cerebellum. Using single-nucleus RNA sequencing in mice, chickens, and humans, as well as STARmap spatial transcriptomic analysis and whole-central nervous system projection tracing, we identified a conserved cell-type set containing two region-specific excitatory neuron classes and three region-invariant inhibitory neuron classes. This set constitutes an archetypal cerebellar nucleus that was repeatedly duplicated to form new regions. The excitatory cell class that preferentially funnels information to lateral frontal cortices in mice becomes predominant in the massively expanded human lateral nucleus. Our data suggest a model of brain region evolution by duplication and divergence of entire cell-type sets.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures


Comment in
-
Adding cognitive connections to the cerebellum.Science. 2020 Dec 18;370(6523):1411-1412. doi: 10.1126/science.abf4483. Science. 2020. PMID: 33335051 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
