

ER-mitochondria contact sites in neurodegeneration: genetic screening approaches to investigate novel disease mechanisms
- PMID: 33335290
- PMCID: PMC8185109
- DOI: 10.1038/s41418-020-00705-8
ER-mitochondria contact sites in neurodegeneration: genetic screening approaches to investigate novel disease mechanisms
Erratum in
-
Correction: ER-mitochondria contact sites in neurodegeneration: genetic screening approaches to investigate novel disease mechanisms.Cell Death Differ. 2021 Oct;28(10):2990. doi: 10.1038/s41418-020-00723-6. Cell Death Differ. 2021. PMID: 33437039 Free PMC article. No abstract available.
Abstract
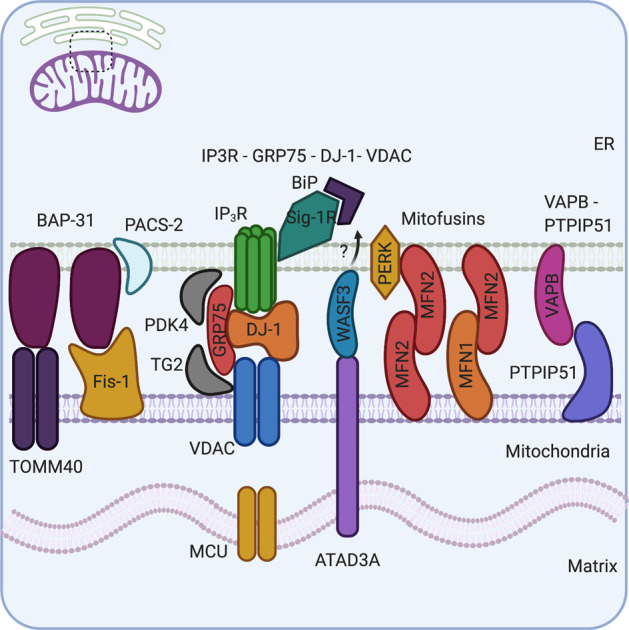
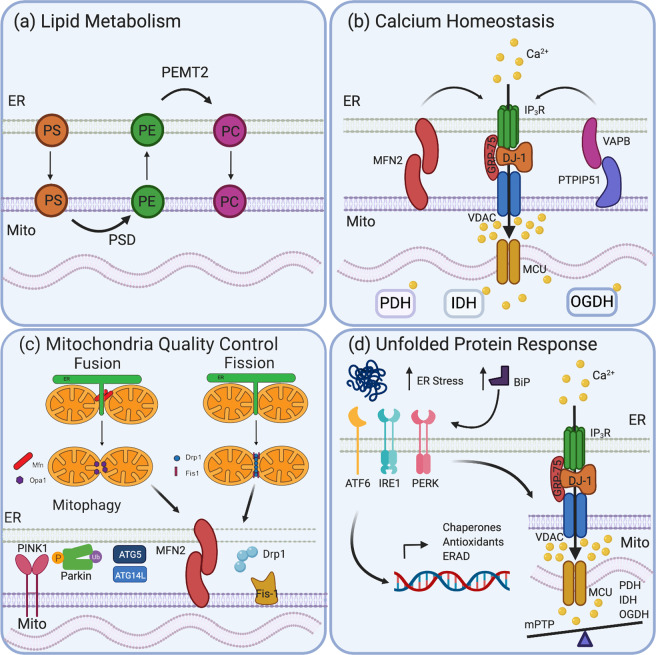
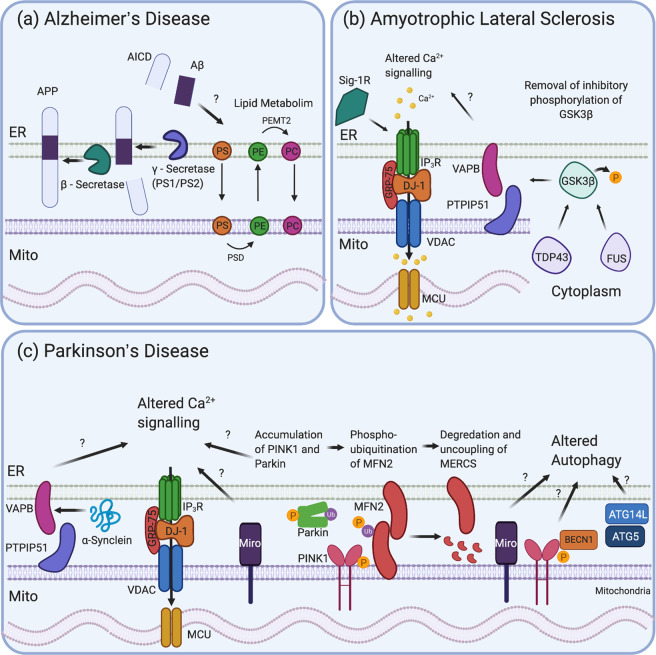
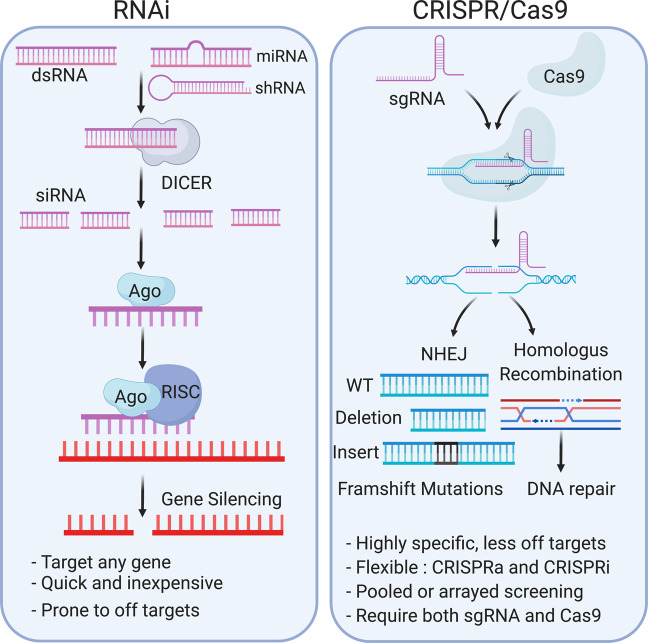
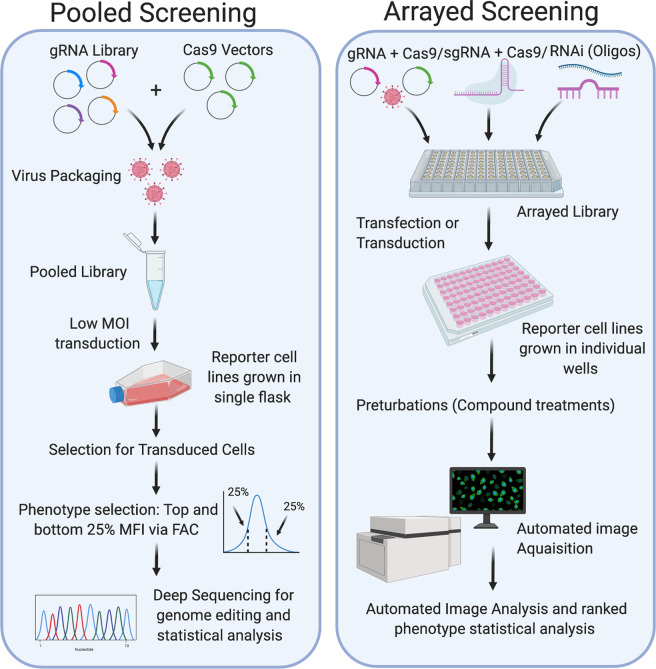
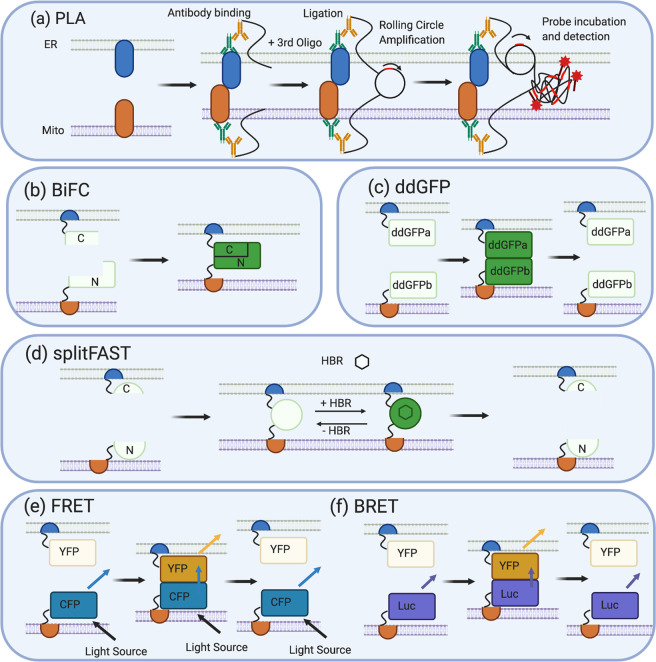
Mitochondria-ER contact sites (MERCS) are known to underpin many important cellular homoeostatic functions, including mitochondrial quality control, lipid metabolism, calcium homoeostasis, the unfolded protein response and ER stress. These functions are known to be dysregulated in neurodegenerative diseases, including Parkinson's disease (PD), Alzheimer's disease (AD) and amyloid lateral sclerosis (ALS), and the number of disease-related proteins and genes being associated with MERCS is increasing. However, many details regarding MERCS and their role in neurodegenerative diseases remain unknown. In this review, we aim to summarise the current knowledge regarding the structure and function of MERCS, and to update the field on current research in PD, AD and ALS. Furthermore, we will evaluate high-throughput screening techniques, including RNAi vs CRISPR/Cas9, pooled vs arrayed formats and how these could be combined with current techniques to visualise MERCS. We will consider the advantages and disadvantages of each technique and how it can be utilised to uncover novel protein pathways involved in MERCS dysfunction in neurodegenerative diseases.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Basso V, Marchesan E, Ziviani E. A trio has turned into a quartet: DJ-1 interacts with the IP3R-Grp75-VDAC complex to control ER-mitochondria interaction. Cell Calcium. 2020. 10.1016/j.ceca.2020.102186. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
