

Secondary metabolites of Bacillus subtilis impact the assembly of soil-derived semisynthetic bacterial communities
- PMID: 33335606
- PMCID: PMC7722629
- DOI: 10.3762/bjoc.16.248
Secondary metabolites of Bacillus subtilis impact the assembly of soil-derived semisynthetic bacterial communities
Abstract
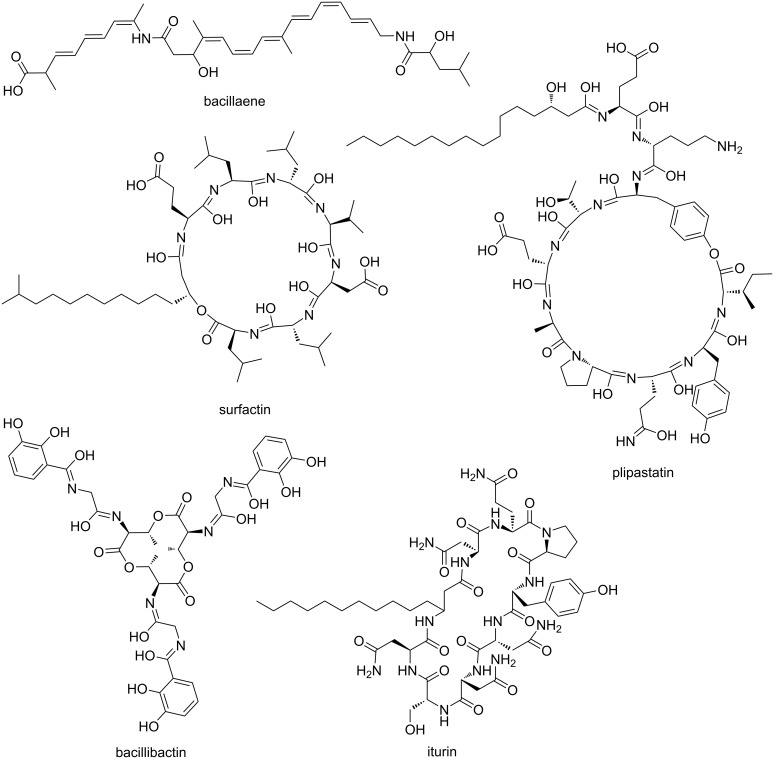
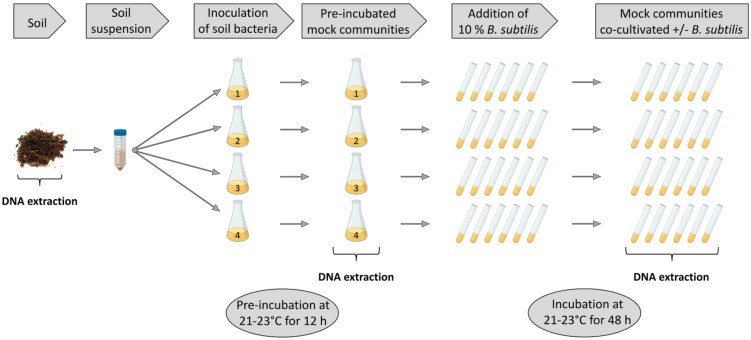
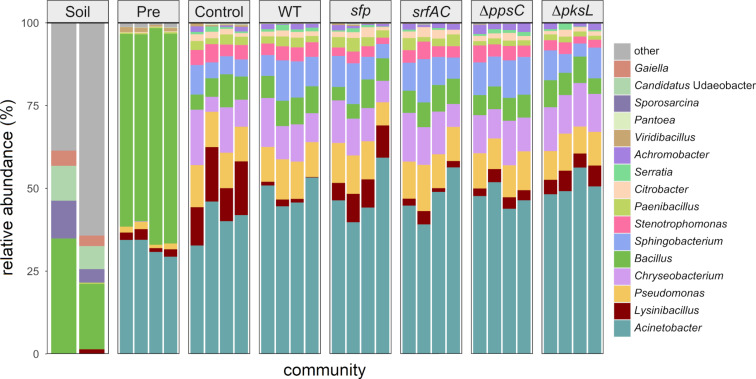
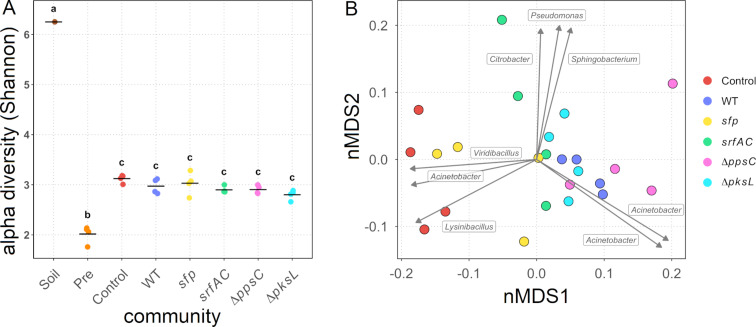
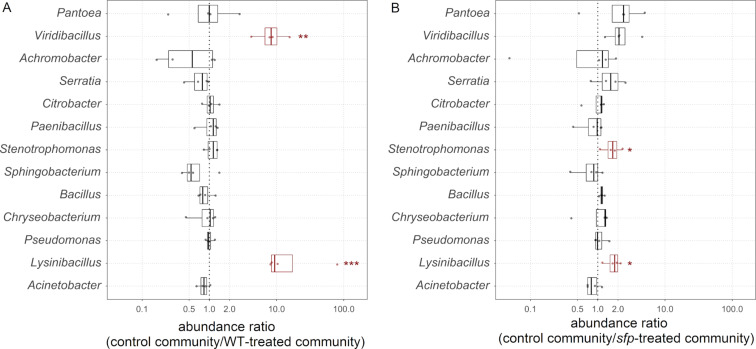
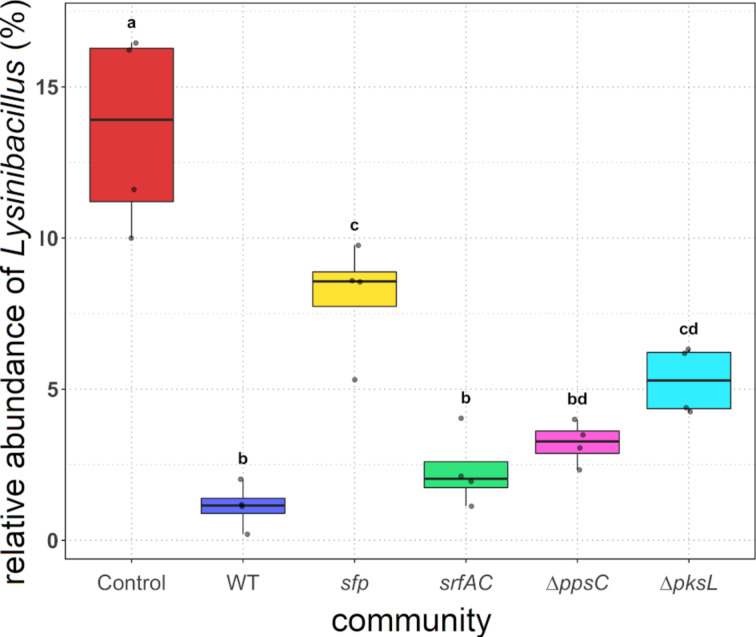
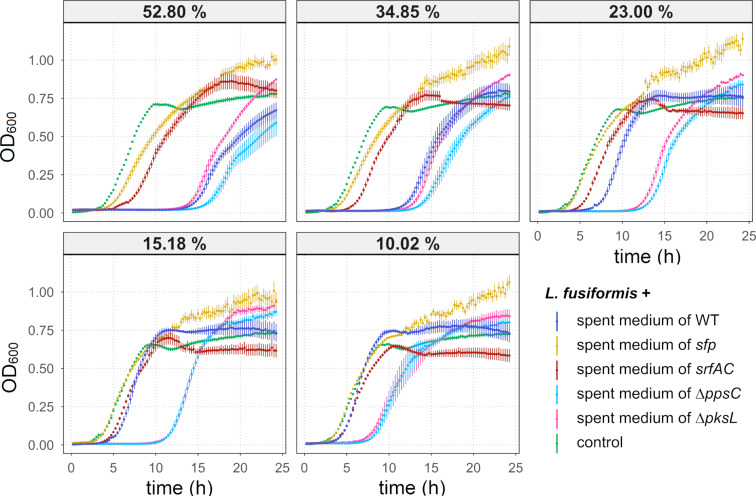
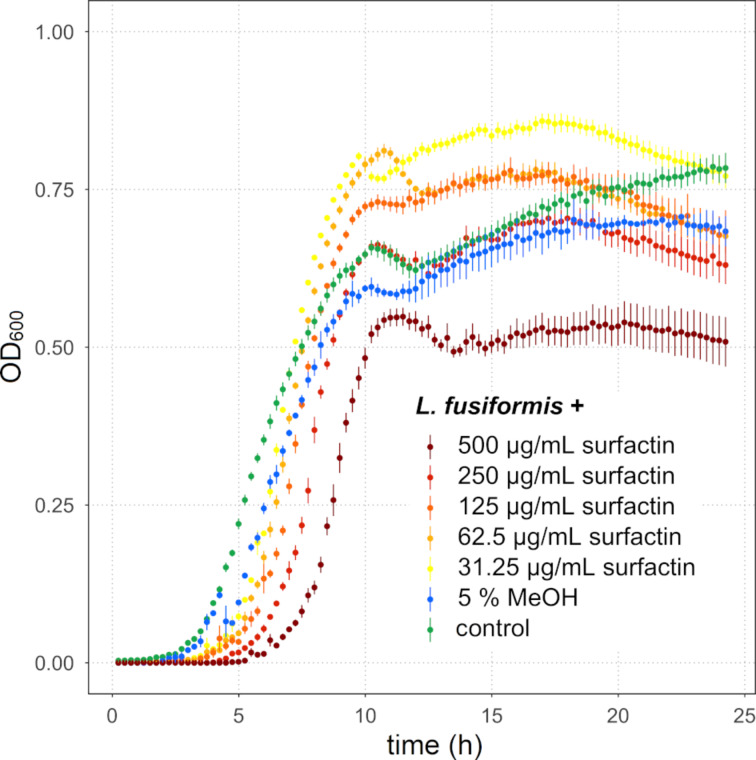
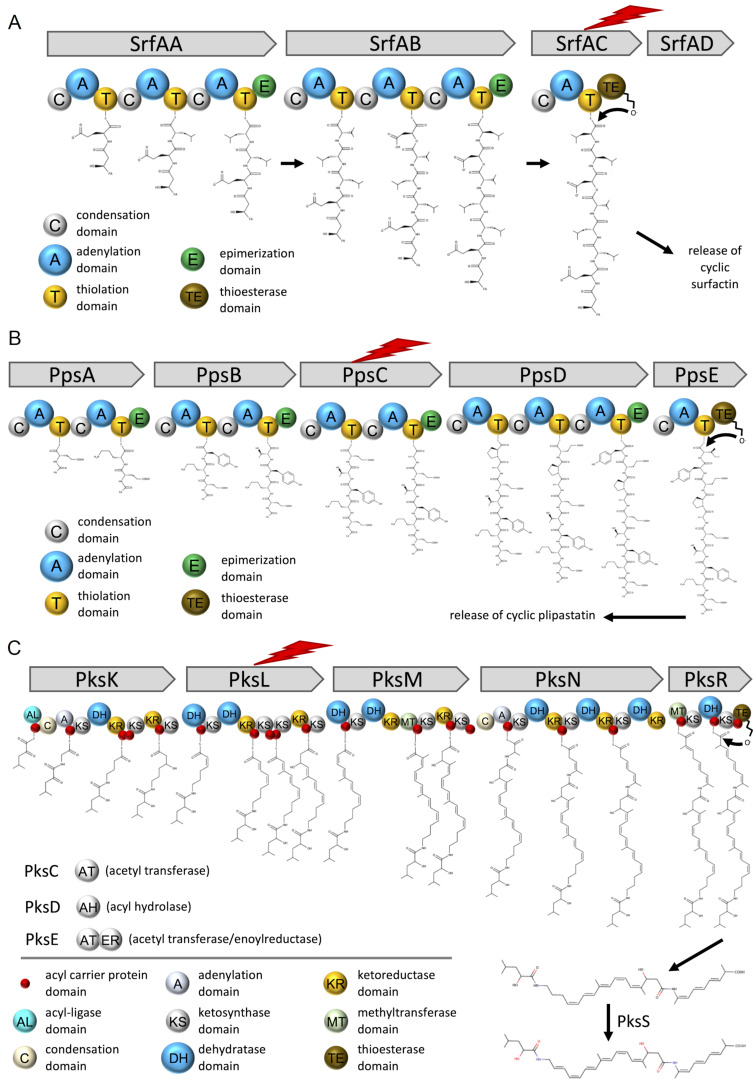
Secondary metabolites provide Bacillus subtilis with increased competitiveness towards other microorganisms. In particular, nonribosomal peptides (NRPs) have an enormous antimicrobial potential by causing cell lysis, perforation of fungal membranes, enzyme inhibition, or disruption of bacterial protein synthesis. This knowledge was primarily acquired in vitro when B. subtilis was competing with other microbial monocultures. However, our understanding of the true ecological role of these small molecules is limited. In this study, we have established soil-derived semisynthetic mock communities containing 13 main genera and supplemented them with B. subtilis P5_B1 WT, the NRP-deficient strain sfp, or single-NRP mutants incapable of producing surfactin, plipastatin, or bacillaene. Through 16S amplicon sequencing, it was revealed that the invasion of NRP-producing B. subtilis strains had no major impact on the bacterial communities. Still, the abundance of the two genera Lysinibacillus and Viridibacillus was reduced. Interestingly, this effect was diminished in communities supplemented with the NRP-deficient strain. Growth profiling of Lysinibacillus fusiformis M5 exposed to either spent media of the B. subtilis strains or pure surfactin indicated the sensitivity of this strain towards the biosurfactant surfactin. Our study provides a more in-depth insight into the influence of B. subtilis NRPs on semisynthetic bacterial communities and helps to understand their ecological role.
Keywords: Bacillus subtilis; Lysinibacillus fusiformis; bacterial community; chemical ecology; nonribosomal peptides; surfactin.
Copyright © 2020, Kiesewalter et al.; licensee Beilstein-Institut.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous