

Metabolic substrate utilization in stress-induced immune cells
- PMID: 33336295
- PMCID: PMC7746792
- DOI: 10.1186/s40635-020-00316-0
Metabolic substrate utilization in stress-induced immune cells
Abstract
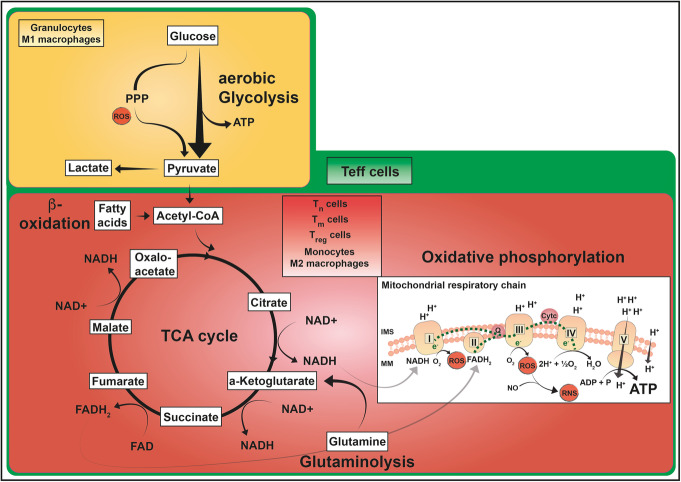
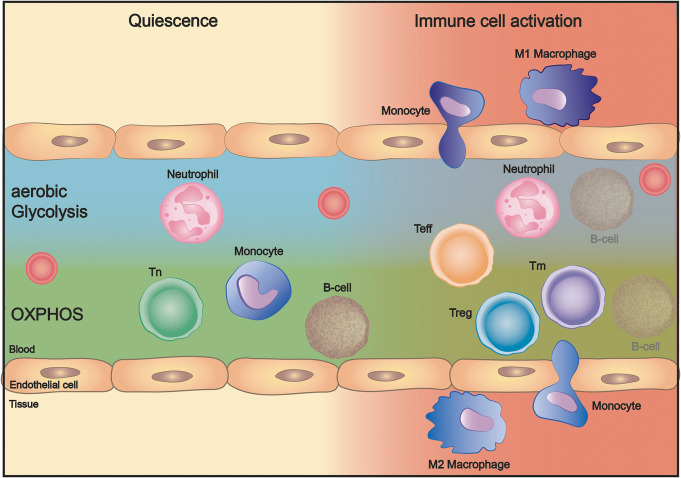
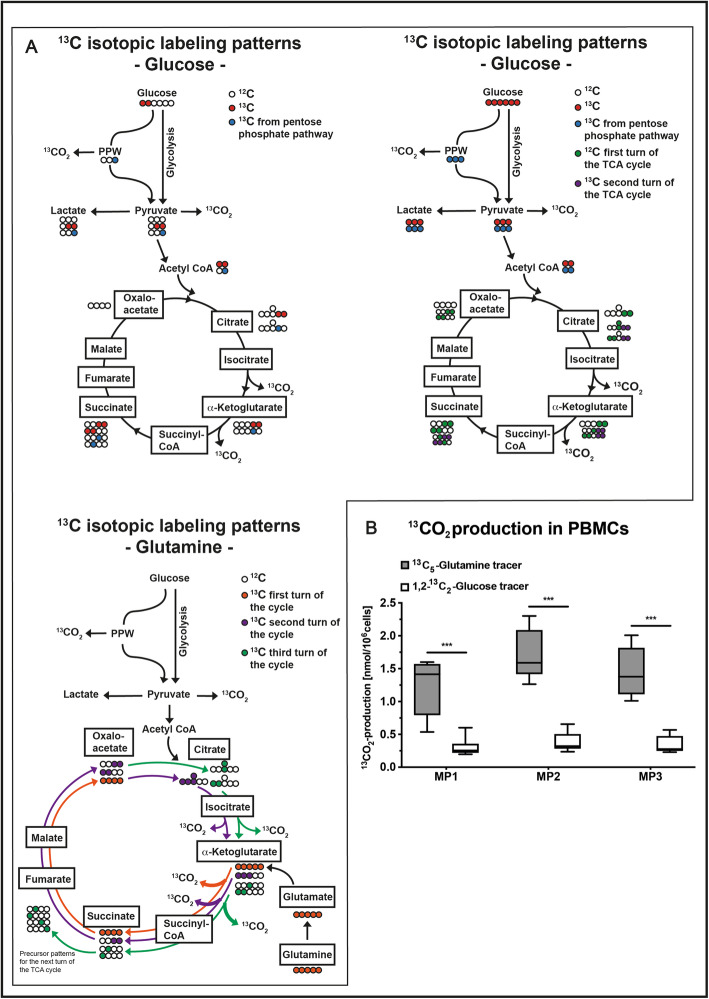
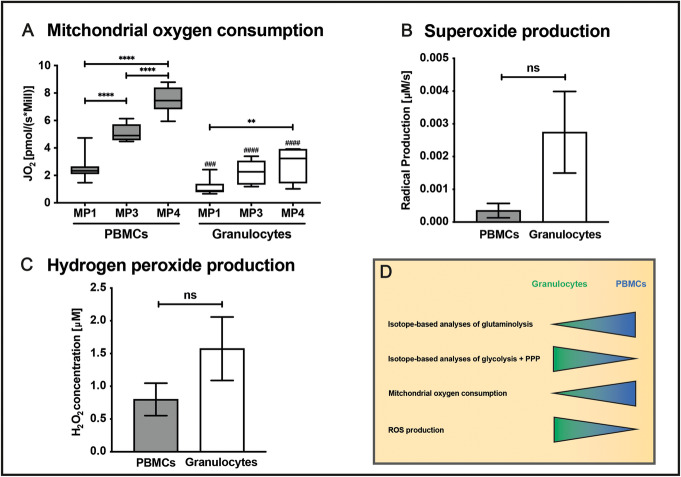
Immune cell activation leads to the acquisition of new functions, such as proliferation, chemotaxis, and cytokine production. These functional changes require continuous metabolic adaption in order to sustain ATP homeostasis for sufficient host defense. The bioenergetic demands are usually met by the interconnected metabolic pathways glycolysis, TCA cycle, and oxidative phosphorylation. Apart from glucose, other sources, such as fatty acids and glutamine, are able to fuel the TCA cycle.Rising evidence has shown that cellular metabolism has a direct effect on the regulation of immune cell functions. Thus, quiescent immune cells maintain a basal metabolic state, which shifts to an accelerated metabolic level upon immune cell activation in order to promote key effector functions.This review article summarizes distinct metabolic signatures of key immune cell subsets from quiescence to activation and demonstrates a methodical concept of how to assess cellular metabolic pathways. It further discusses why metabolic functions are of rising interest for translational research and how they can be affected by the underlying pathophysiological condition and/or therapeutic interventions.
Keywords: Catecholamines; Glycolysis; Immunometabolism; Oxidative phosphorylation; Pentose phosphate pathway; Reactive oxygen species; Tricarboxylic acid cycle.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Warburg O. On the origin of cancer cells. Science (80- ) 1956;123:309–314. - PubMed
-
- Medzhitov R, Janeway C. Innate immune recognition: mechanisms and pathways. Immunol Rev. 2000;173:89–97. - PubMed
-
- Maianski NA, Maianski AN, Kuijpers TW, Roos D. Apoptosis of Neutrophils. Acta Haematol. 2004;111:56–66. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
