

Inflammasome activation and evasion by bacterial pathogens
- PMID: 33338767
- PMCID: PMC7925435
- DOI: 10.1016/j.coi.2020.11.006
Inflammasome activation and evasion by bacterial pathogens
Abstract
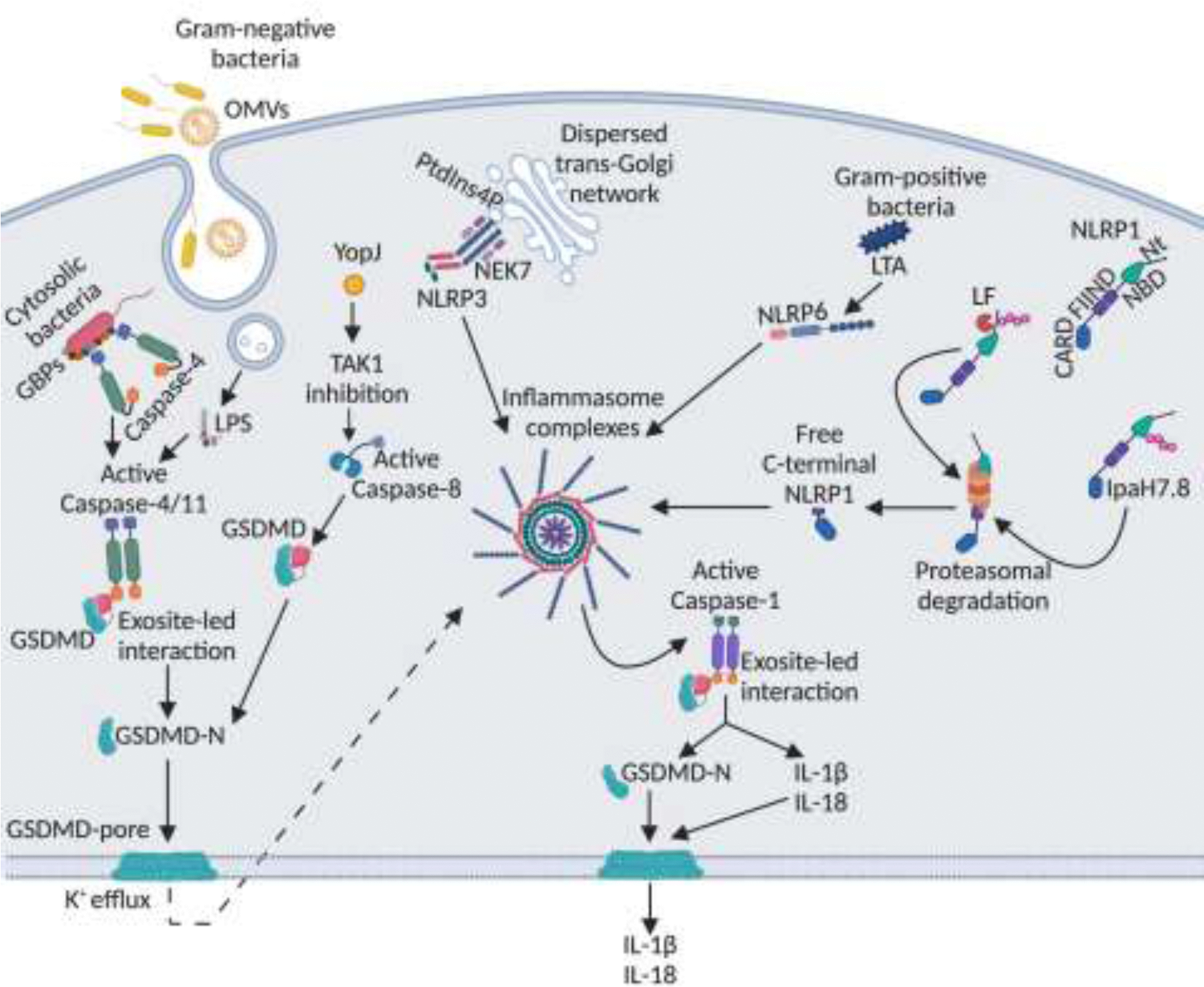
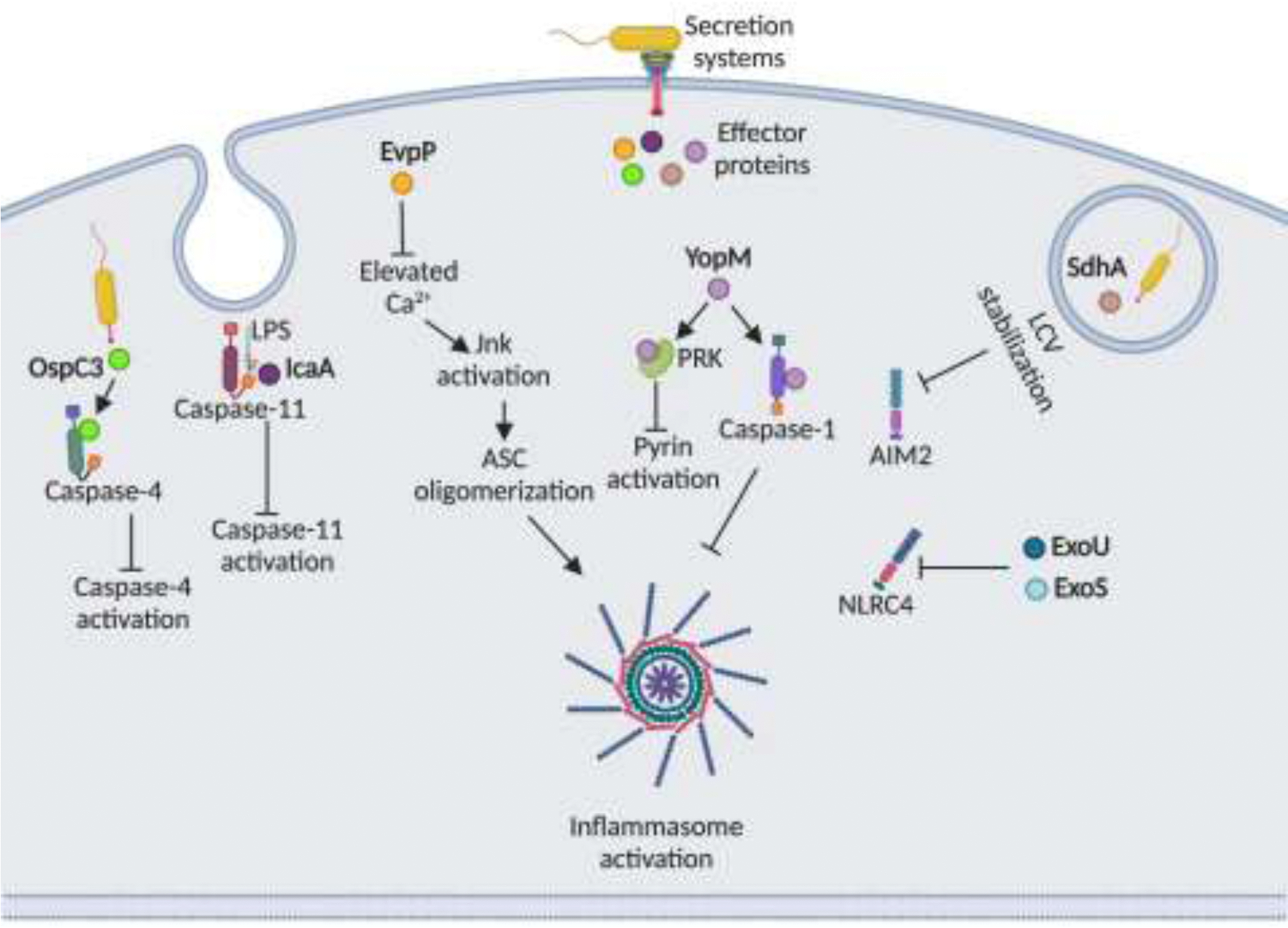
Innate immune system plays an essential role in combating infectious diseases by recognizing invading pathogens and activating host defense response. Inflammasomes complexes are a central component of the cytosolic innate immune surveillance and are vital in host defense against bacterial pathogens. Bacterial products or pathogen-induced modifications in the intracellular environment are sensed by the inflammasome receptors that form complexes that serve as a platform for caspase-1-dependent or caspase-11-dependent induction of pyroptosis and secretion of cytokines, IL-1β and IL-18. However, several pathogenic bacteria have developed strategies to evade inflammasome activation. This review highlights the recent advances in the mechanism of inflammasome activation by bacterial pathogens and some of the bacterial evasion strategies of inflammasome activation.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
References
-
- Martinon F, Burns K, Tschopp J: The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Molecular cell 2002, 10:417 426. - PubMed
-
- Shi J, Zhao Y, Wang K, Shi X, Wang Y, Huang H, Zhuang Y, Cai T, Wang F, Shao F: Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526:660–665. - PubMed
-
- Kayagaki N, Stowe IB, Lee BL, O’Rourke K, Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT, et al. : Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526:666–671. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
