

The SARS-CoV-2 Spike protein has a broad tropism for mammalian ACE2 proteins
- PMID: 33347434
- PMCID: PMC7751883
- DOI: 10.1371/journal.pbio.3001016
The SARS-CoV-2 Spike protein has a broad tropism for mammalian ACE2 proteins
Abstract
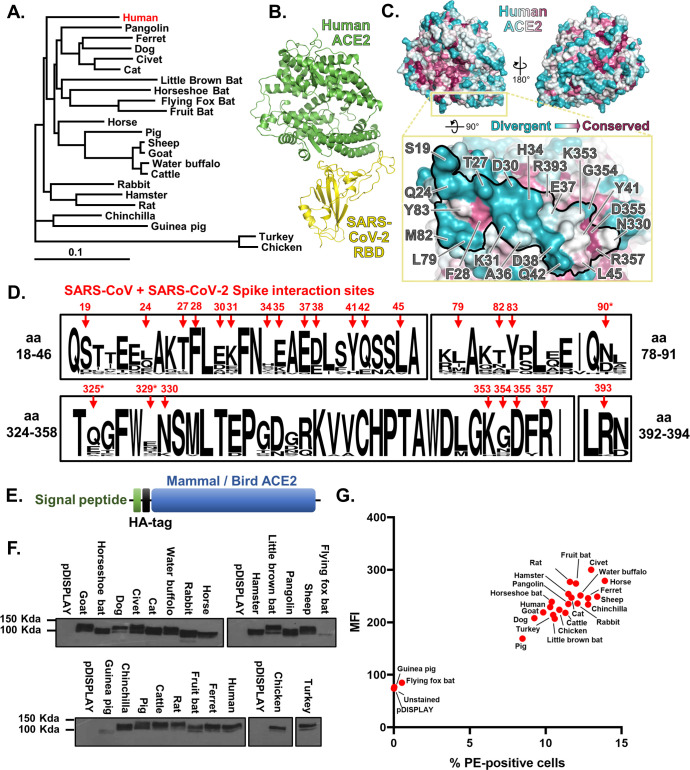
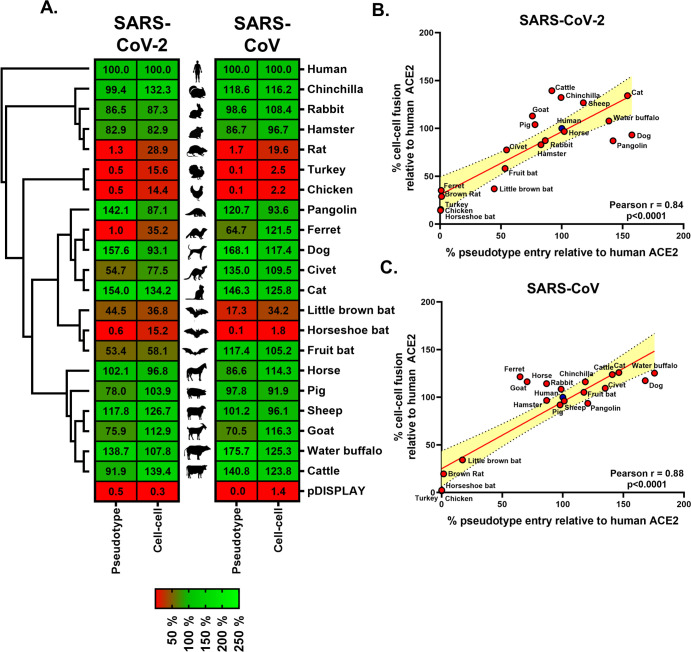
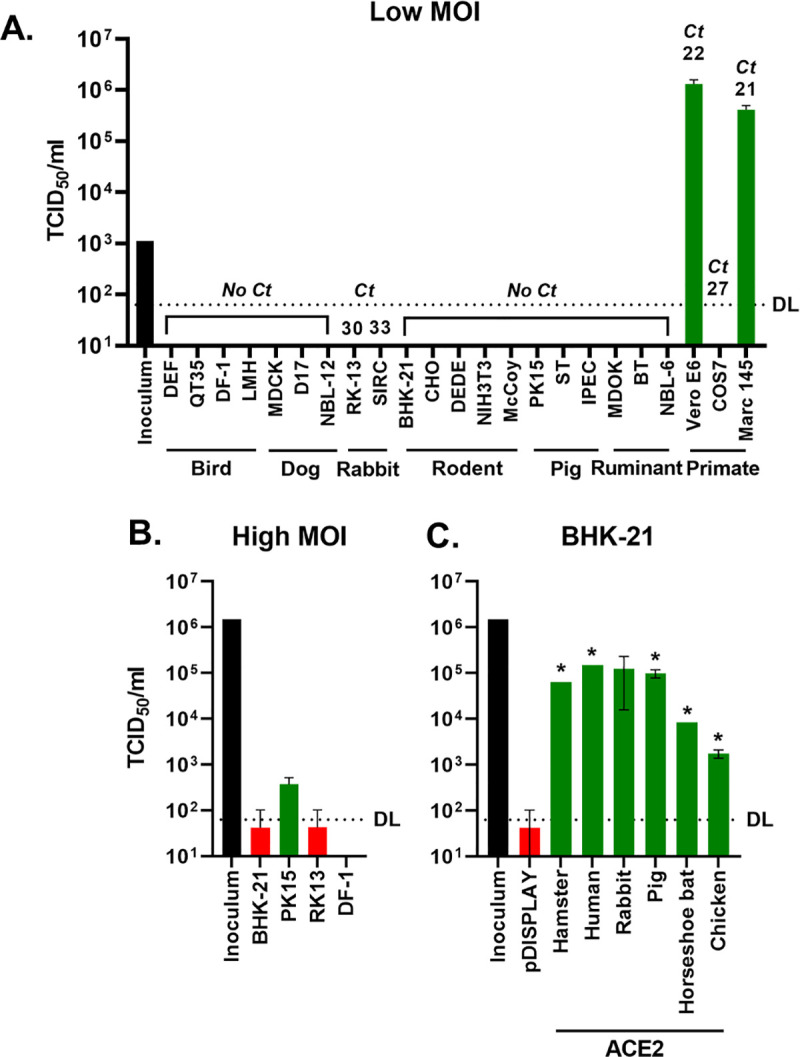
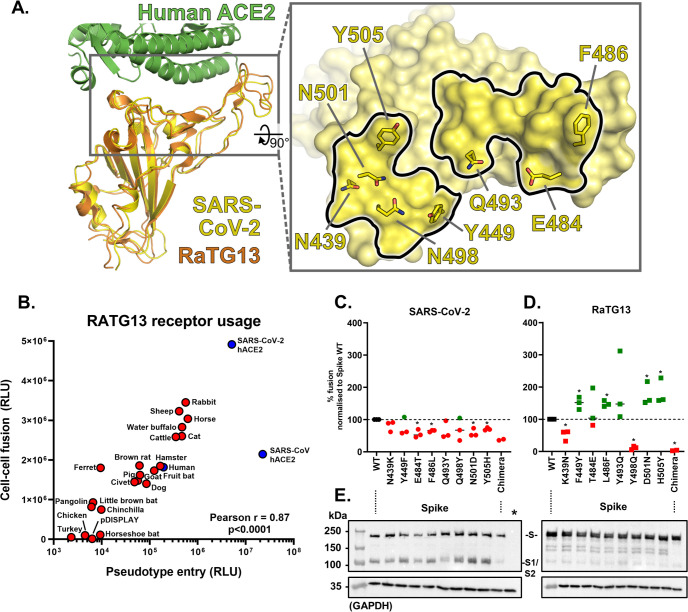
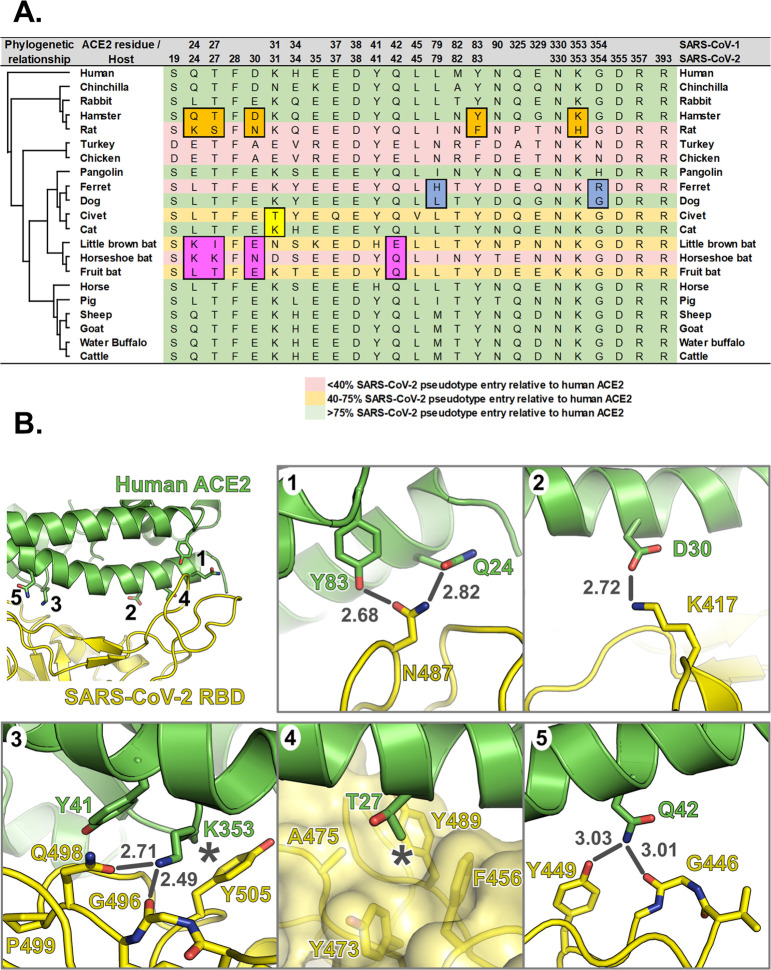
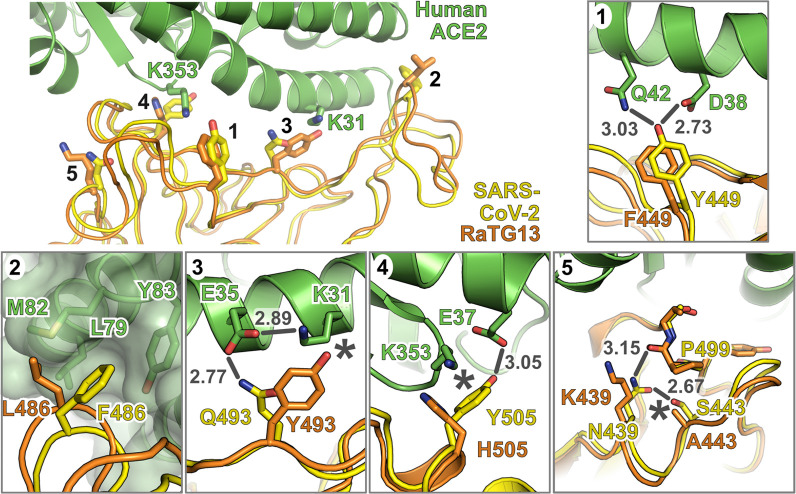
SARS Coronavirus 2 (SARS-CoV-2) emerged in late 2019, leading to the Coronavirus Disease 2019 (COVID-19) pandemic that continues to cause significant global mortality in human populations. Given its sequence similarity to SARS-CoV, as well as related coronaviruses circulating in bats, SARS-CoV-2 is thought to have originated in Chiroptera species in China. However, whether the virus spread directly to humans or through an intermediate host is currently unclear, as is the potential for this virus to infect companion animals, livestock, and wildlife that could act as viral reservoirs. Using a combination of surrogate entry assays and live virus, we demonstrate that, in addition to human angiotensin-converting enzyme 2 (ACE2), the Spike glycoprotein of SARS-CoV-2 has a broad host tropism for mammalian ACE2 receptors, despite divergence in the amino acids at the Spike receptor binding site on these proteins. Of the 22 different hosts we investigated, ACE2 proteins from dog, cat, and cattle were the most permissive to SARS-CoV-2, while bat and bird ACE2 proteins were the least efficiently used receptors. The absence of a significant tropism for any of the 3 genetically distinct bat ACE2 proteins we examined indicates that SARS-CoV-2 receptor usage likely shifted during zoonotic transmission from bats into people, possibly in an intermediate reservoir. Comparison of SARS-CoV-2 receptor usage to the related coronaviruses SARS-CoV and RaTG13 identified distinct tropisms, with the 2 human viruses being more closely aligned. Finally, using bioinformatics, structural data, and targeted mutagenesis, we identified amino acid residues within the Spike-ACE2 interface, which may have played a pivotal role in the emergence of SARS-CoV-2 in humans. The apparently broad tropism of SARS-CoV-2 at the point of viral entry confirms the potential risk of infection to a wide range of companion animals, livestock, and wildlife.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Huang A.T., Garcia-Carreras B., Hitchings M.D.T., Yang B., Katzelnick L., Rattigan S.M., Borgert B., Moreno C., Solomon B.D., Rodriguez-Barraquer I., Lessler J., Salje H., Burke D.S., Wesolowski A., and Cummings D.A.T., A systematic review of antibody mediated immunity to coronaviruses: antibody kinetics, correlates of protection, and association of antibody responses with severity of disease. medRxiv, 2020. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BBS/E/I/00007034/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00007037/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 098406/Z/12/B/WT_/Wellcome Trust/United Kingdom
- BBS/E/I/COV07001/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/P021735/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
