

Dopamine as a Multifunctional Neurotransmitter in Gastropod Molluscs: An Evolutionary Hypothesis
- PMID: 33347799
- PMCID: PMC8016498
- DOI: 10.1086/711293
Dopamine as a Multifunctional Neurotransmitter in Gastropod Molluscs: An Evolutionary Hypothesis
Abstract
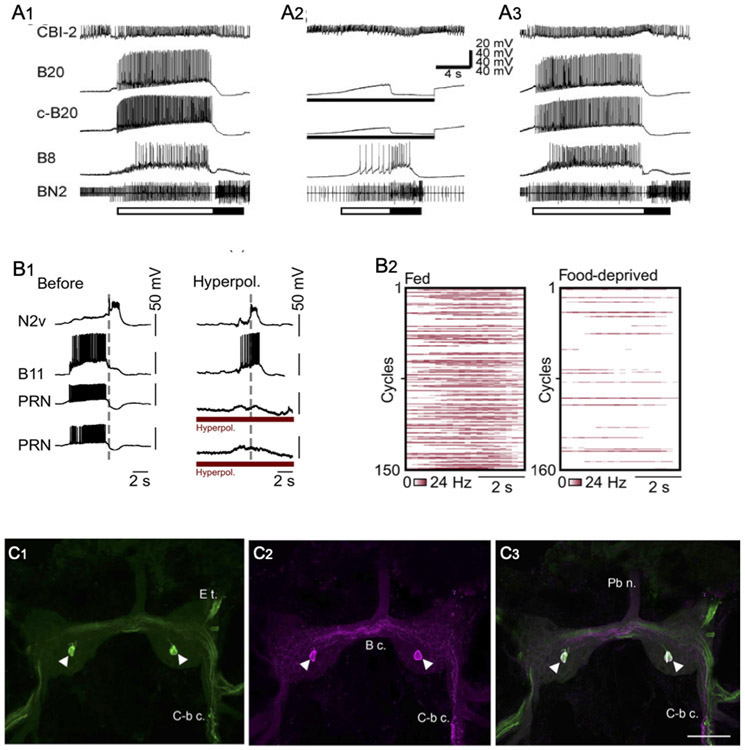
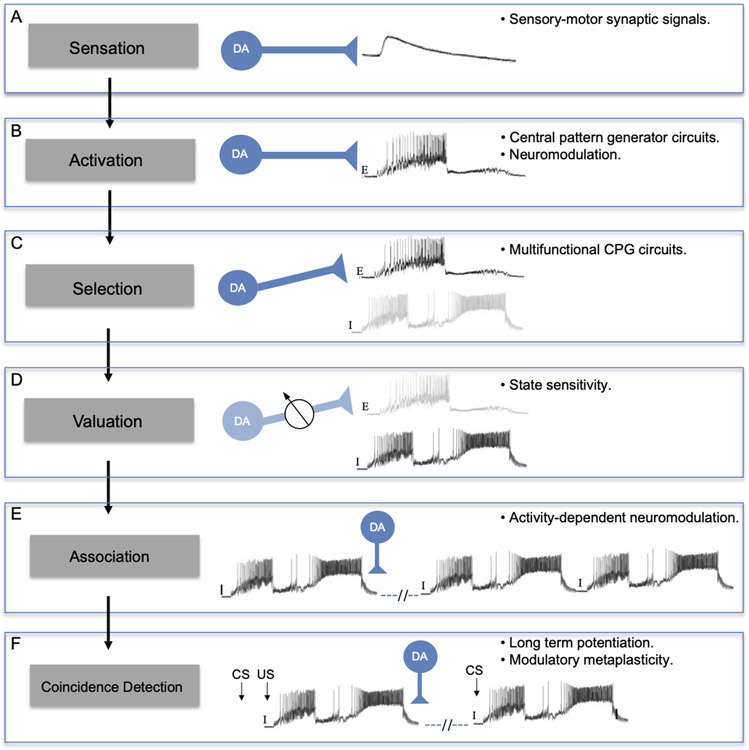
AbstractThe catecholamine 3,4-dihydroxyphenethylamine, or dopamine, acts as a neurotransmitter across a broad phylogenetic spectrum. Functions attributed to dopamine in the mammalian brain include regulation of motor circuits, valuation of sensory stimuli, and mediation of reward or reinforcement signals. Considerable evidence also supports a neurotransmitter role for dopamine in gastropod molluscs, and there is growing appreciation for its potential common functions across phylogeny. This article reviews evidence for dopamine's transmitter role in the nervous systems of gastropods. The functional properties of identified dopaminergic neurons in well-characterized neural circuits suggest a hypothetical incremental sequence by which dopamine accumulated its diverse roles. The successive acquisition of dopamine functions is proposed in the context of gastropod feeding behavior: (1) sensation of potential nutrients, (2) activation of motor circuits, (3) selection of motor patterns from multifunctional circuits, (4) valuation of sensory stimuli with reference to internal state, (5) association of motor programs with their outcomes, and (6) coincidence detection between sensory stimuli and their consequences. At each stage of this sequence, it is proposed that existing functions of dopaminergic neurons favored their recruitment to fulfill additional information processing demands. Common functions of dopamine in other intensively studied groups, ranging from mammals and insects to nematodes, suggest an ancient origin for this progression.
Figures
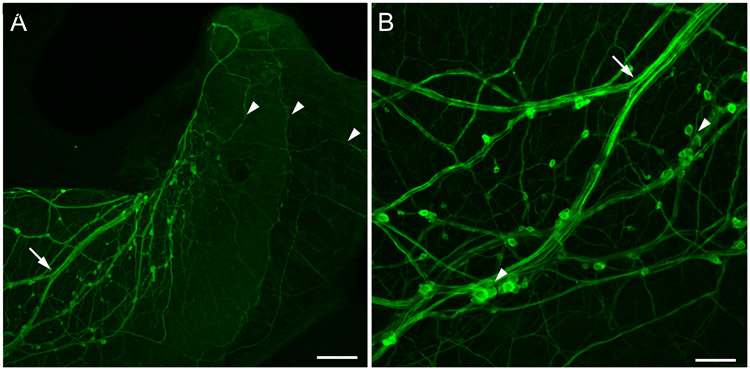
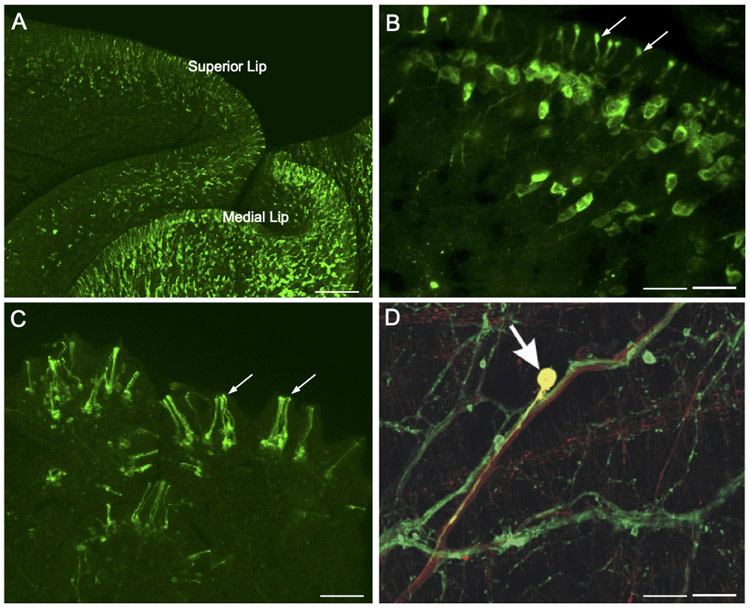
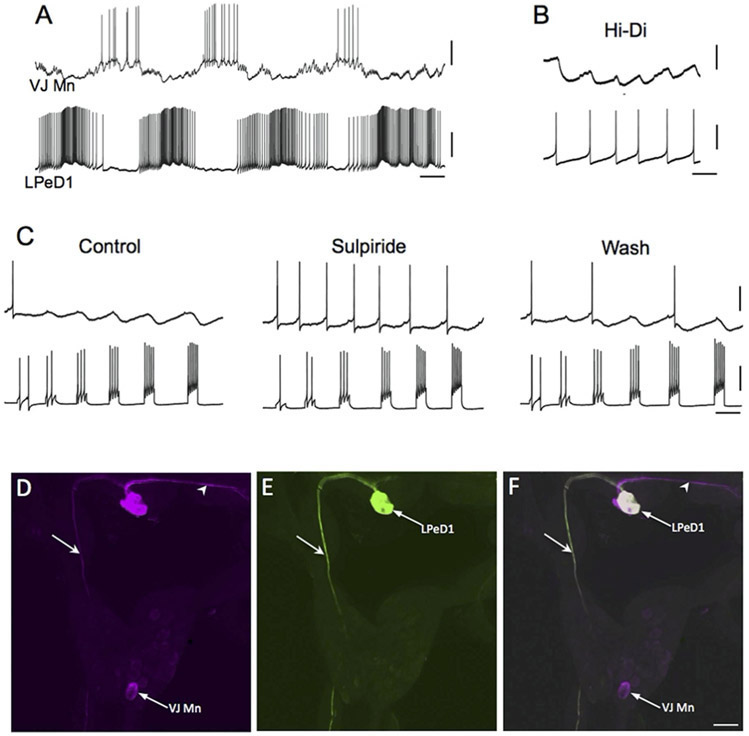
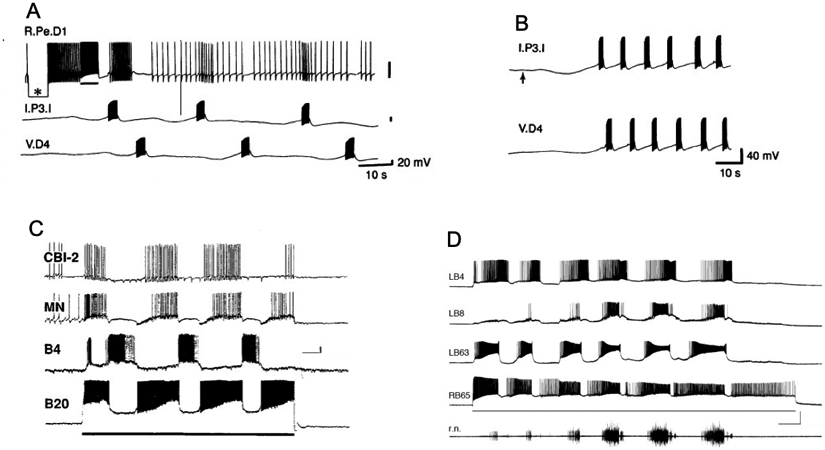
Similar articles
-
GABA as a Neurotransmitter in Gastropod Molluscs.Biol Bull. 2019 Apr;236(2):144-156. doi: 10.1086/701377. Epub 2019 Jan 16. Biol Bull. 2019. PMID: 30933636 Free PMC article. Review.
-
Dopaminergic Central Neurons and Peripheral Sensory Systems in Pteropod and Nudibranch Molluscs.J Comp Neurol. 2025 May;533(5):e70054. doi: 10.1002/cne.70054. J Comp Neurol. 2025. PMID: 40287934
-
The importance of identified neurons in gastropod molluscs to neuroscience.Curr Opin Neurobiol. 2019 Jun;56:1-7. doi: 10.1016/j.conb.2018.10.009. Epub 2018 Nov 1. Curr Opin Neurobiol. 2019. PMID: 30390485 Review.
-
Discrete neurochemical coding of distinguishable motivational processes: insights from nucleus accumbens control of feeding.Psychopharmacology (Berl). 2007 Apr;191(3):439-59. doi: 10.1007/s00213-007-0741-z. Epub 2007 Feb 23. Psychopharmacology (Berl). 2007. PMID: 17318502 Review.
-
The debate over dopamine's role in reward: the case for incentive salience.Psychopharmacology (Berl). 2007 Apr;191(3):391-431. doi: 10.1007/s00213-006-0578-x. Epub 2006 Oct 27. Psychopharmacology (Berl). 2007. PMID: 17072591 Review.
Cited by
-
The Gut-Brain Axis in Schizophrenia: The Implications of the Gut Microbiome and SCFA Production.Nutrients. 2023 Oct 16;15(20):4391. doi: 10.3390/nu15204391. Nutrients. 2023. PMID: 37892465 Free PMC article. Review.
-
Neurotransmitter metabolites in milk ferments of Leuconostoc mesenteroides regulate temperature-sensitive heartbeats in an ex ovo model.Heliyon. 2024 Aug 11;10(16):e36129. doi: 10.1016/j.heliyon.2024.e36129. eCollection 2024 Aug 30. Heliyon. 2024. PMID: 39253113 Free PMC article.
-
Application of neurotransmitters and dental stem cells for pulp regeneration: A review.Saudi Dent J. 2023 Jul;35(5):387-394. doi: 10.1016/j.sdentj.2023.05.004. Epub 2023 May 12. Saudi Dent J. 2023. PMID: 37520592 Free PMC article. Review.
-
Melatonin's Impact on Cytokine Storm and Modulation of Purinergic Receptors for COVID-19 Prognosis: A Mental Health Perspective.J Mol Neurosci. 2024 Dec 5;74(4):113. doi: 10.1007/s12031-024-02292-6. J Mol Neurosci. 2024. PMID: 39636363 Review.
-
A peripheral subepithelial network for chemotactile processing in the predatory sea slug Pleurobranchaea californica.PLoS One. 2024 Feb 8;19(2):e0296872. doi: 10.1371/journal.pone.0296872. eCollection 2024. PLoS One. 2024. PMID: 38329975 Free PMC article.
References
-
- Anderson WW, and Barker DL. 1981. Synaptic mechanisms that generate network oscillations in the absence of discrete synaptic potentials. J. Exp. Zool 216:187–191. - PubMed
-
- Ascher P 1968. Electrophoretic injections of dopamine on Aplysia neurones. J. Physiol 198: 48–49 P.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
