

A ubiquitin switch controls autocatalytic inactivation of the DNA-protein crosslink repair protease SPRTN
- PMID: 33348378
- PMCID: PMC7826251
- DOI: 10.1093/nar/gkaa1224
A ubiquitin switch controls autocatalytic inactivation of the DNA-protein crosslink repair protease SPRTN
Abstract
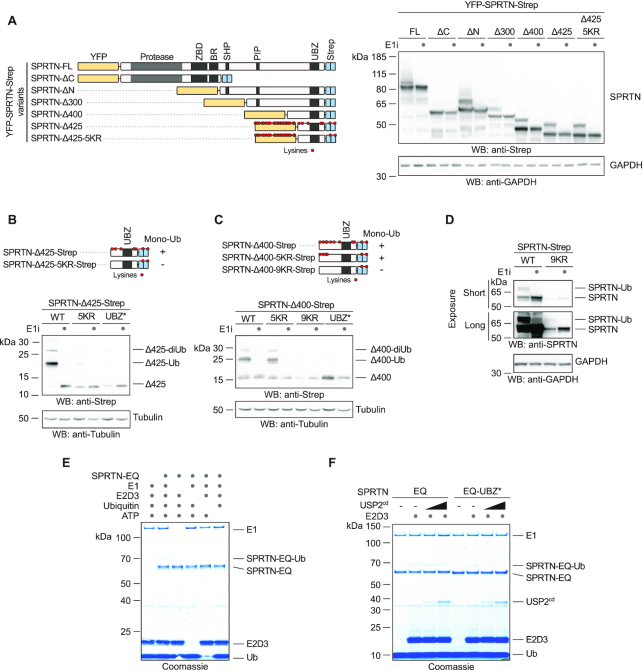
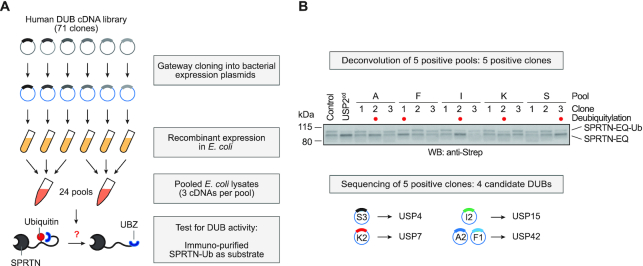
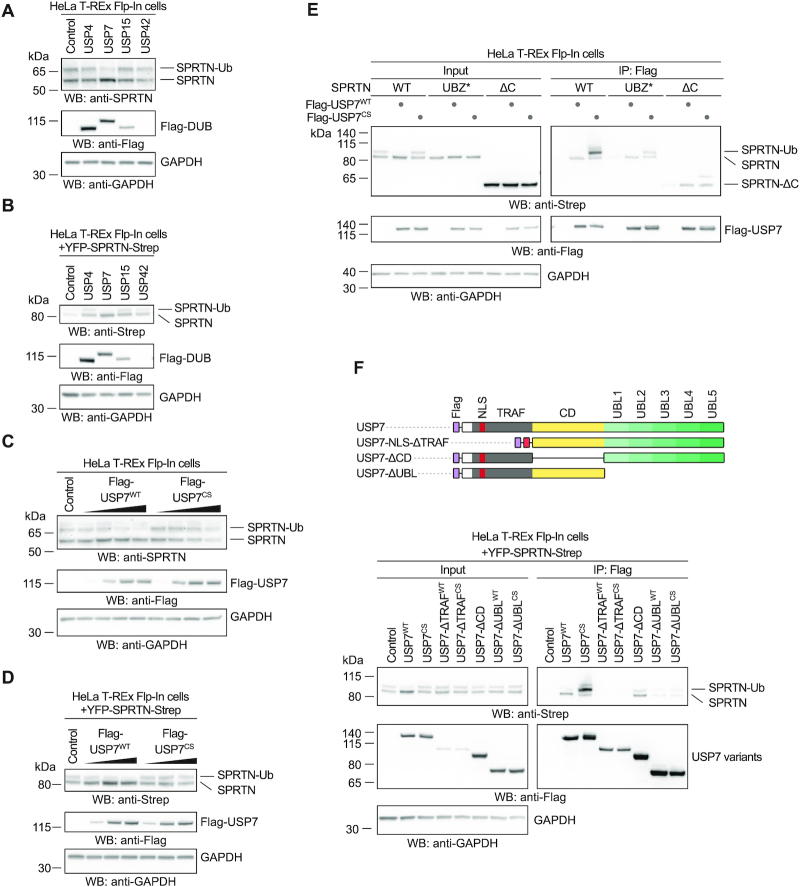
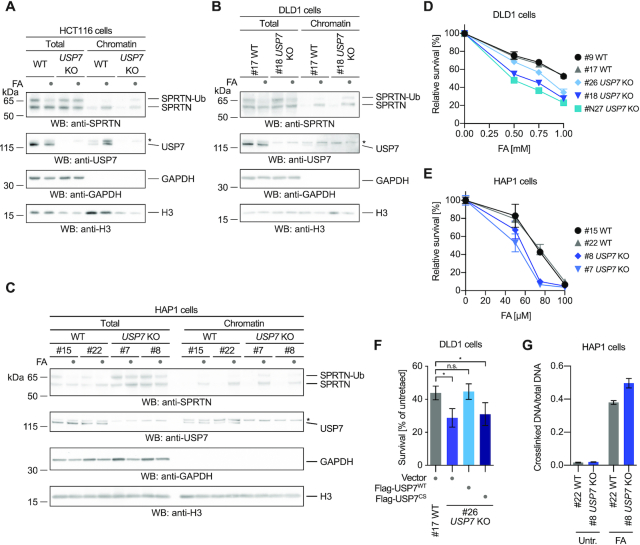
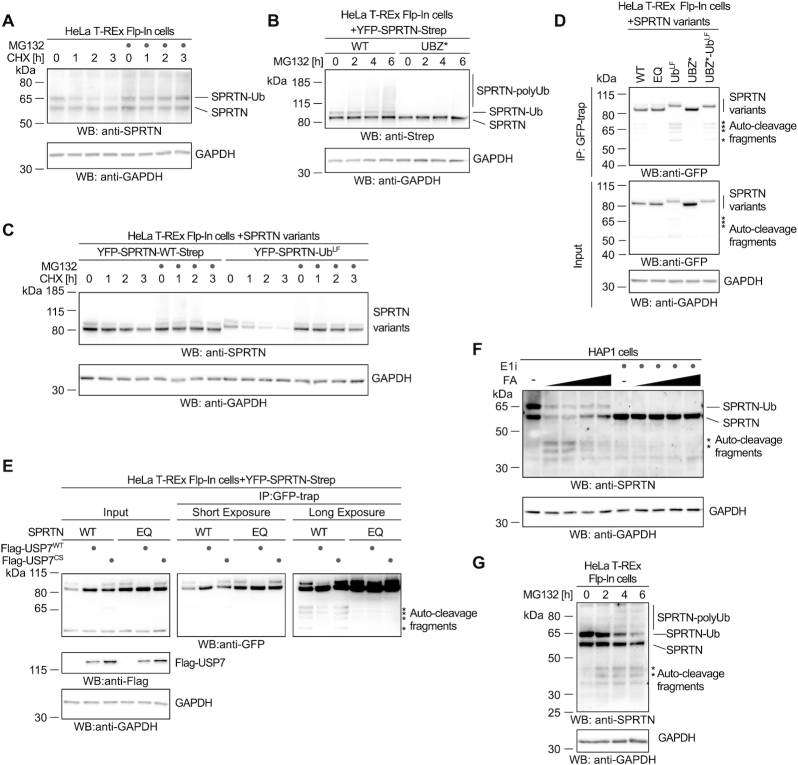
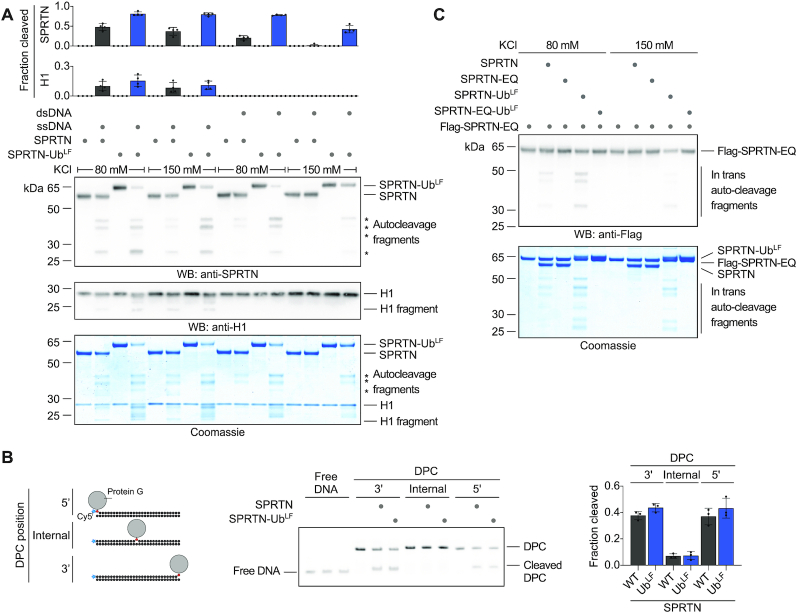
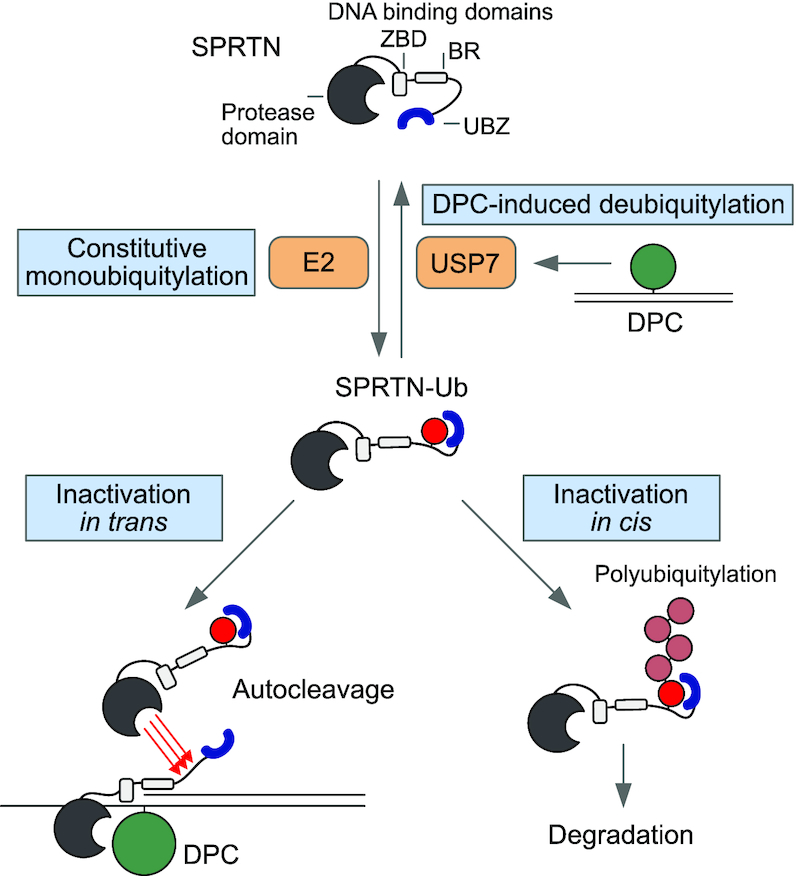
Repair of covalent DNA-protein crosslinks (DPCs) by the metalloprotease SPRTN prevents genome instability, premature aging and carcinogenesis. SPRTN is specifically activated by DNA structures containing single- and double-stranded features, but degrades the protein components of DPCs promiscuously and independent of amino acid sequence. This lack of specificity is useful to target diverse protein adducts, however, it requires tight control in return, in order to prohibit uncontrolled proteolysis of chromatin proteins. Here, we discover the components and principles of a ubiquitin switch, which negatively regulates SPRTN. We demonstrate that monoubiquitylation is induced in an E3 ligase-independent manner and, in contrast to previous assumptions, does not control chromatin access of the enzyme. Data obtained in cells and in vitro reveal that monoubiquitylation induces inactivation of the enzyme by triggering autocatalytic cleavage in trans while also priming SPRTN for proteasomal degradation in cis. Finally, we show that the deubiquitylating enzyme USP7 antagonizes this negative control of SPRTN in the presence of DPCs.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
