

The Impact of the 'Mis-Peptidome' on HLA Class I-Mediated Diseases: Contribution of ERAP1 and ERAP2 and Effects on the Immune Response
- PMID: 33348540
- PMCID: PMC7765998
- DOI: 10.3390/ijms21249608
The Impact of the 'Mis-Peptidome' on HLA Class I-Mediated Diseases: Contribution of ERAP1 and ERAP2 and Effects on the Immune Response
Abstract
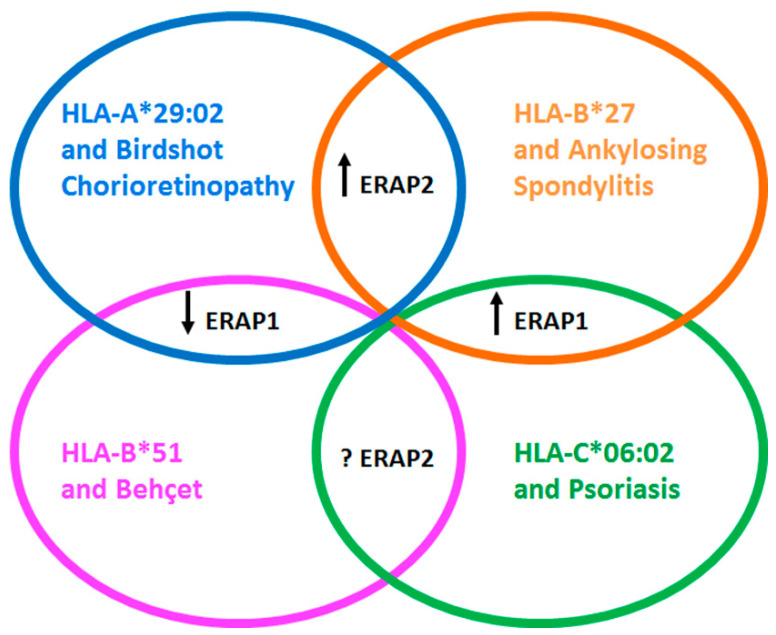
The strong association with the Major Histocompatibility Complex (MHC) class I genes represents a shared trait for a group of autoimmune/autoinflammatory disorders having in common immunopathogenetic basis as well as clinical features. Accordingly, the main risk factors for Ankylosing Spondylitis (AS), prototype of the Spondyloarthropathies (SpA), the Behçet's disease (BD), the Psoriasis (Ps) and the Birdshot Chorioretinopathy (BSCR) are HLA-B*27, HLA-B*51, HLA-C*06:02 and HLA-A*29:02, respectively. Despite the strength of the association, the HLA pathogenetic role in these diseases is far from being thoroughly understood. Furthermore, Genome-Wide Association Studies (GWAS) have highlighted other important susceptibility factors such as Endoplasmic Reticulum Aminopeptidase (ERAP) 1 and, less frequently, ERAP2 that refine the peptidome presented by HLA class I molecules to CD8+ T cells. Mass spectrometry analysis provided considerable knowledge of HLA-B*27, HLA-B*51, HLA-C*06:02 and HLA-A*29:02 immunopeptidome. However, the combined effect of several ERAP1 and ERAP2 allelic variants could generate an altered pool of peptides accounting for the "mis-immunopeptidome" that ranges from suboptimal to pathogenetic/harmful peptides able to induce non-canonical or autoreactive CD8+ T responses, activation of NK cells and/or garbling the classical functions of the HLA class I molecules. This review will focus on this class of epitopes as possible elicitors of atypical/harmful immune responses which can contribute to the pathogenesis of chronic inflammatory diseases.
Keywords: CD8+ T cells; ERAP1 and ERAP2 ER aminopeptidases; HLA class I molecules; antigen presentation; autoimmune/autoinflammatory disorders; immunopeptidome.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
