

Enriched conditioning expands the regenerative ability of sensory neurons after spinal cord injury via neuronal intrinsic redox signaling
- PMID: 33349630
- PMCID: PMC7752916
- DOI: 10.1038/s41467-020-20179-z
Enriched conditioning expands the regenerative ability of sensory neurons after spinal cord injury via neuronal intrinsic redox signaling
Abstract
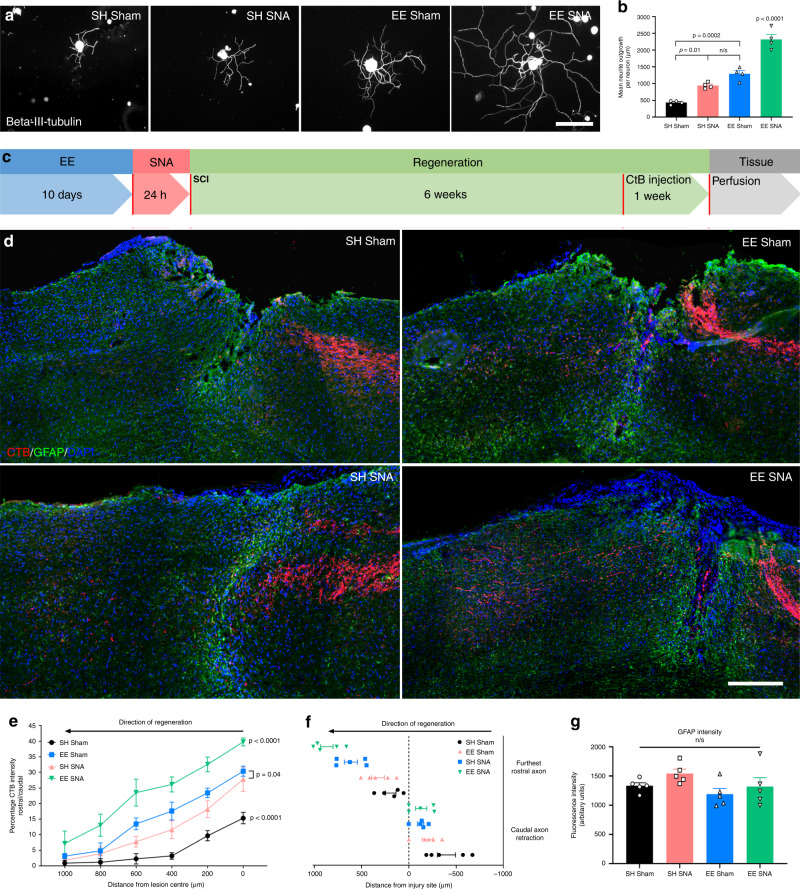
Overcoming the restricted axonal regenerative ability that limits functional repair following a central nervous system injury remains a challenge. Here we report a regenerative paradigm that we call enriched conditioning, which combines environmental enrichment (EE) followed by a conditioning sciatic nerve axotomy that precedes a spinal cord injury (SCI). Enriched conditioning significantly increases the regenerative ability of dorsal root ganglia (DRG) sensory neurons compared to EE or a conditioning injury alone, propelling axon growth well beyond the spinal injury site. Mechanistically, we established that enriched conditioning relies on the unique neuronal intrinsic signaling axis PKC-STAT3-NADPH oxidase 2 (NOX2), enhancing redox signaling as shown by redox proteomics in DRG. Finally, NOX2 conditional deletion or overexpression respectively blocked or phenocopied enriched conditioning-dependent axon regeneration after SCI leading to improved functional recovery. These studies provide a paradigm that drives the regenerative ability of sensory neurons offering a potential redox-dependent regenerative model for mechanistic and therapeutic discoveries.
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
AMPK controls the axonal regenerative ability of dorsal root ganglia sensory neurons after spinal cord injury.Nat Metab. 2020 Sep;2(9):918-933. doi: 10.1038/s42255-020-0252-3. Epub 2020 Aug 10. Nat Metab. 2020. PMID: 32778834
-
Rapamycin-Resistant mTOR Activity Is Required for Sensory Axon Regeneration Induced by a Conditioning Lesion.eNeuro. 2017 Jan 13;3(6):ENEURO.0358-16.2016. doi: 10.1523/ENEURO.0358-16.2016. eCollection 2016 Nov-Dec. eNeuro. 2017. PMID: 28101526 Free PMC article.
-
Reactive oxygen species regulate axonal regeneration through the release of exosomal NADPH oxidase 2 complexes into injured axons.Nat Cell Biol. 2018 Mar;20(3):307-319. doi: 10.1038/s41556-018-0039-x. Epub 2018 Feb 12. Nat Cell Biol. 2018. PMID: 29434374
-
The Dorsal Column Lesion Model of Spinal Cord Injury and Its Use in Deciphering the Neuron-Intrinsic Injury Response.Dev Neurobiol. 2018 Oct;78(10):926-951. doi: 10.1002/dneu.22601. Epub 2018 May 11. Dev Neurobiol. 2018. PMID: 29717546 Free PMC article. Review.
-
Differential regenerative ability of sensory and motor neurons.Neurosci Lett. 2017 Jun 23;652:35-40. doi: 10.1016/j.neulet.2016.11.004. Epub 2016 Nov 3. Neurosci Lett. 2017. PMID: 27818349 Review.
Cited by
-
CBP/p300 activation promotes axon growth, sprouting, and synaptic plasticity in chronic experimental spinal cord injury with severe disability.PLoS Biol. 2022 Sep 20;20(9):e3001310. doi: 10.1371/journal.pbio.3001310. eCollection 2022 Sep. PLoS Biol. 2022. PMID: 36126035 Free PMC article.
-
Molecular networking and collision cross section prediction for structural isomer and unknown compound identification in plant metabolomics: a case study applied to Zhanthoxylum heitzii extracts.Anal Bioanal Chem. 2022 Jun;414(14):4103-4118. doi: 10.1007/s00216-022-04059-7. Epub 2022 Apr 13. Anal Bioanal Chem. 2022. PMID: 35419692
-
Discovery of therapeutic targets for spinal cord injury based on molecular mechanisms of axon regeneration after conditioning lesion.J Transl Med. 2023 Jul 28;21(1):511. doi: 10.1186/s12967-023-04375-1. J Transl Med. 2023. PMID: 37507810 Free PMC article.
-
Exploring marine-derived bacterial compounds targeting the μ-opioid receptor agonists through metabolic profiling to molecular modeling.Sci Rep. 2025 May 17;15(1):17196. doi: 10.1038/s41598-025-96411-x. Sci Rep. 2025. PMID: 40382432 Free PMC article.
-
Dorsal Root Injury-A Model for Exploring Pathophysiology and Therapeutic Strategies in Spinal Cord Injury.Cells. 2021 Aug 25;10(9):2185. doi: 10.3390/cells10092185. Cells. 2021. PMID: 34571835 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
