

A selective transmembrane recognition mechanism by a membrane-anchored ubiquitin ligase adaptor
- PMID: 33351099
- PMCID: PMC7759299
- DOI: 10.1083/jcb.202001116
A selective transmembrane recognition mechanism by a membrane-anchored ubiquitin ligase adaptor
Abstract
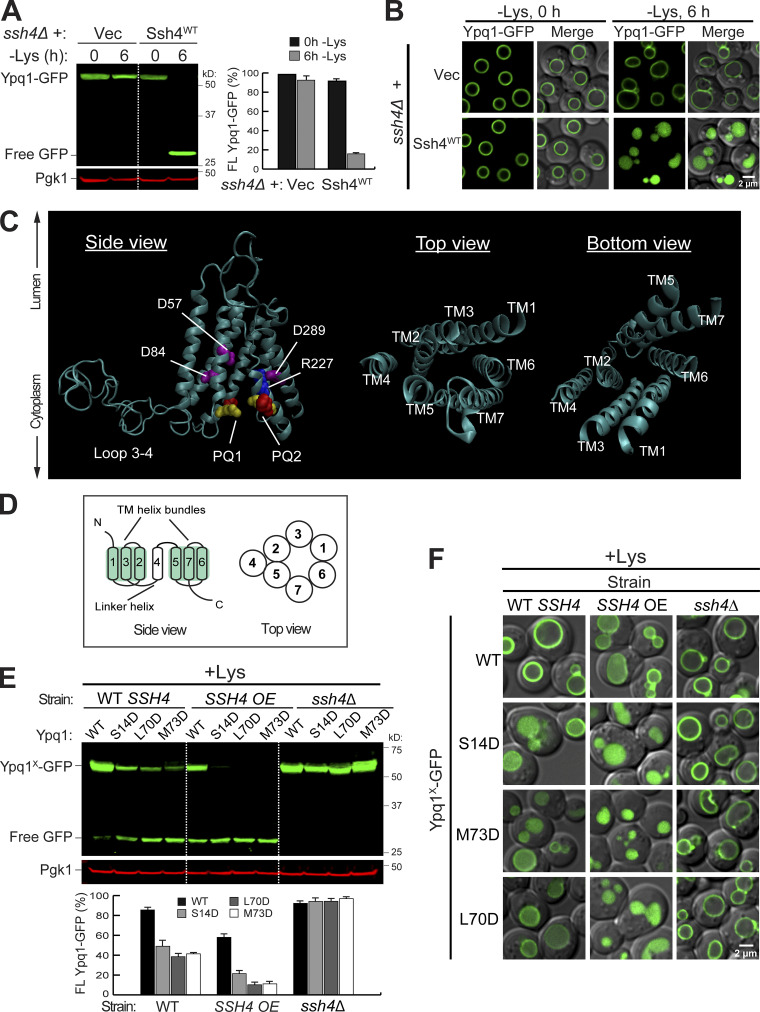
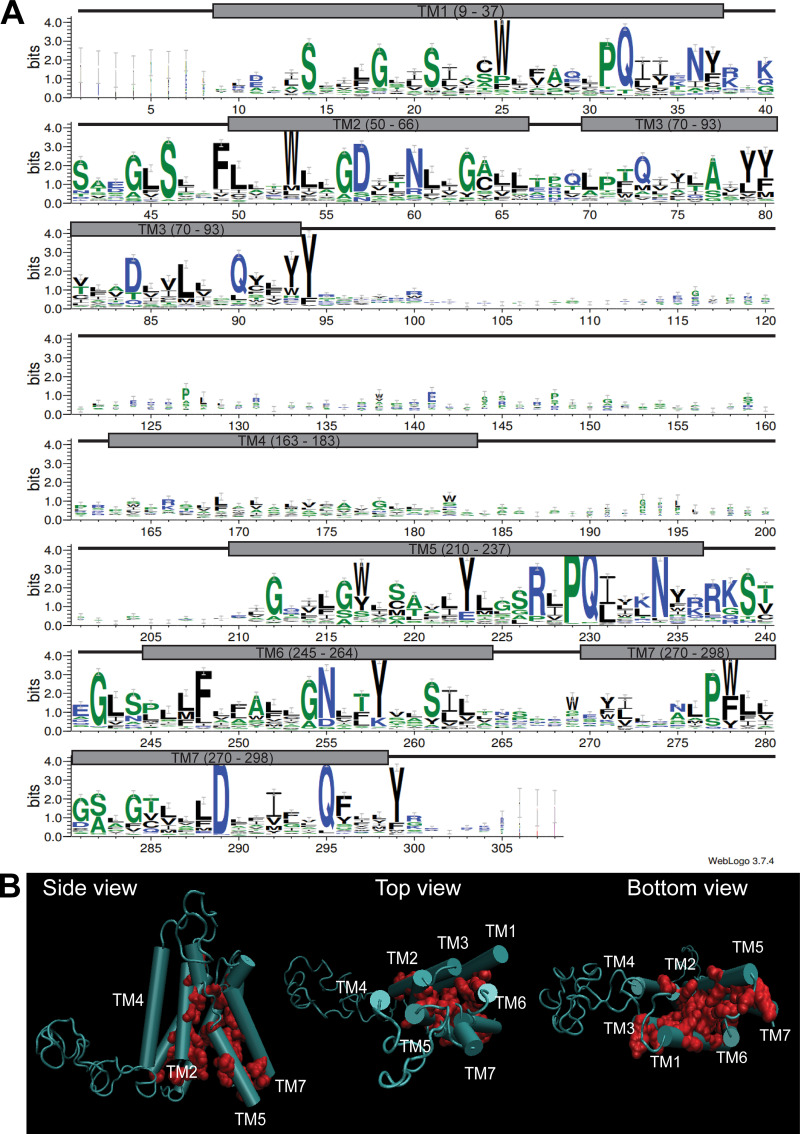
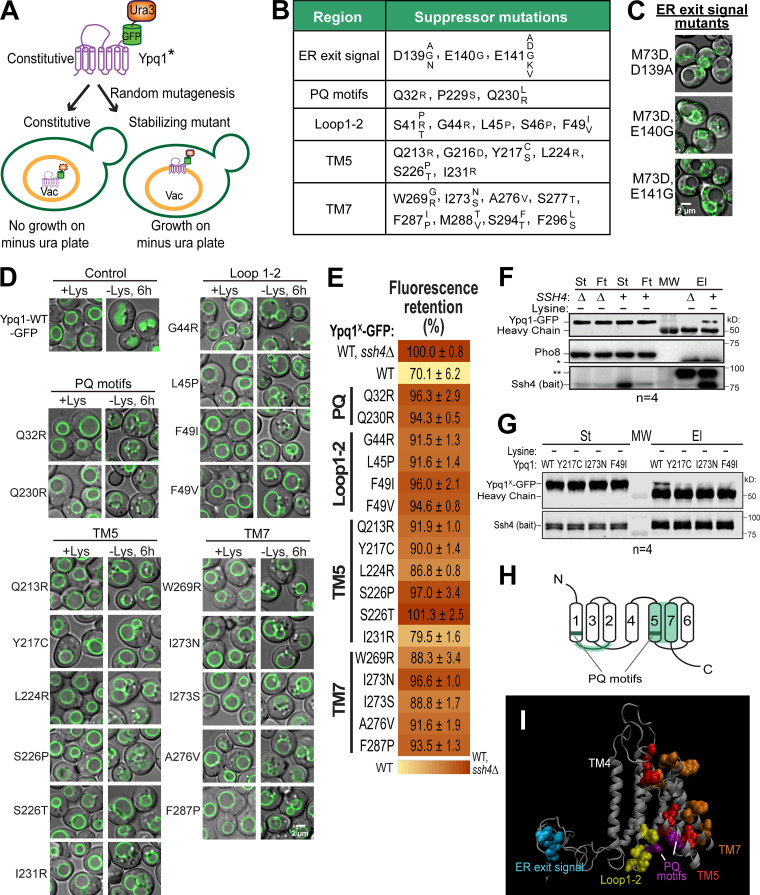
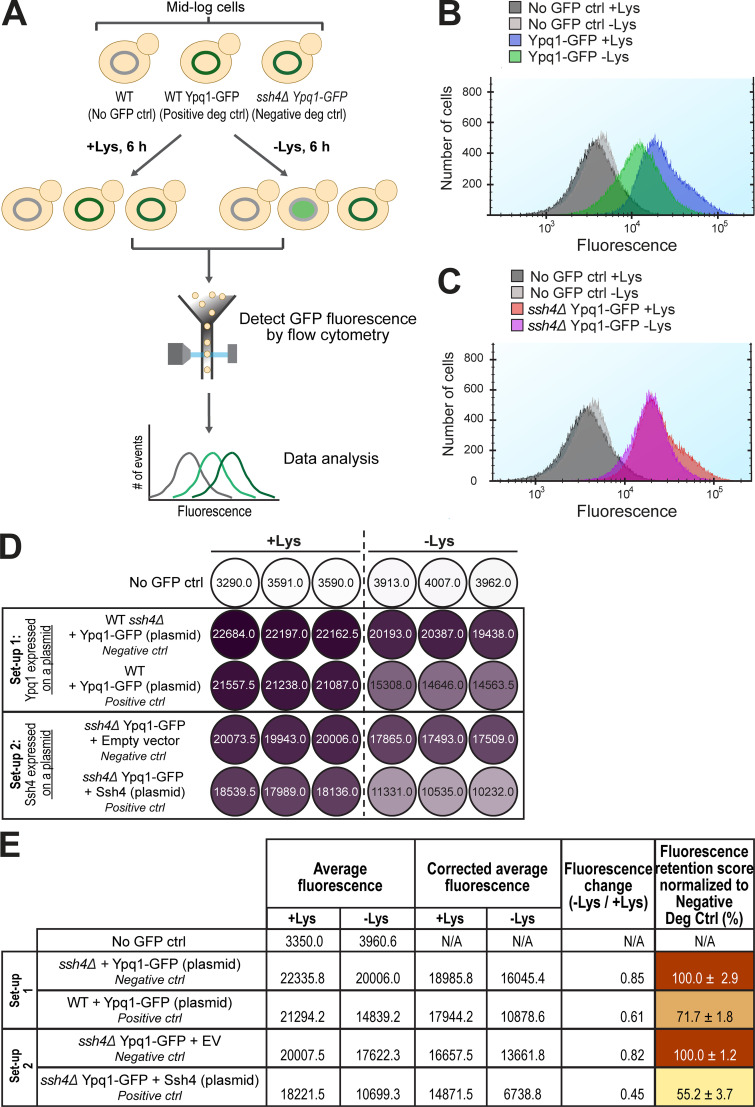
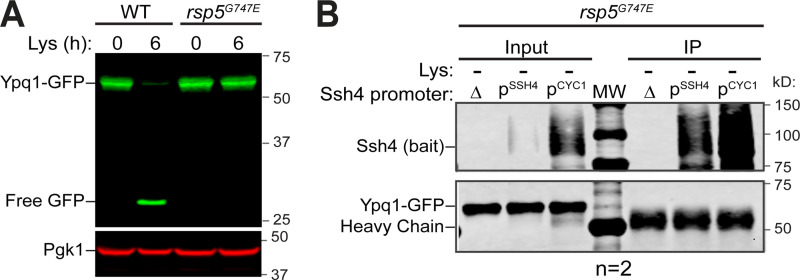
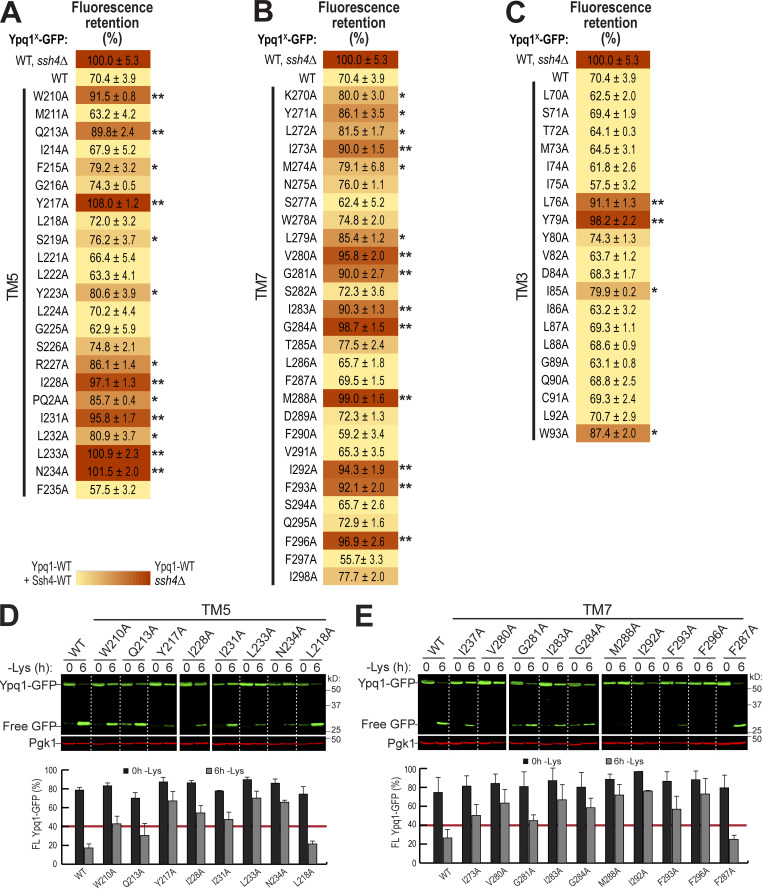
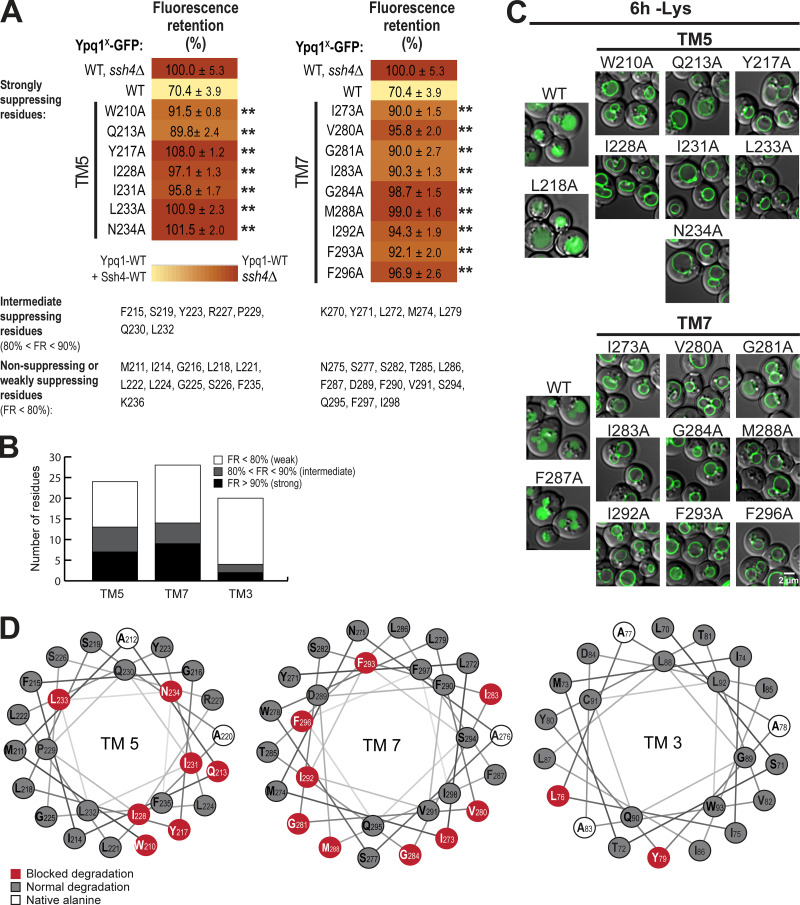
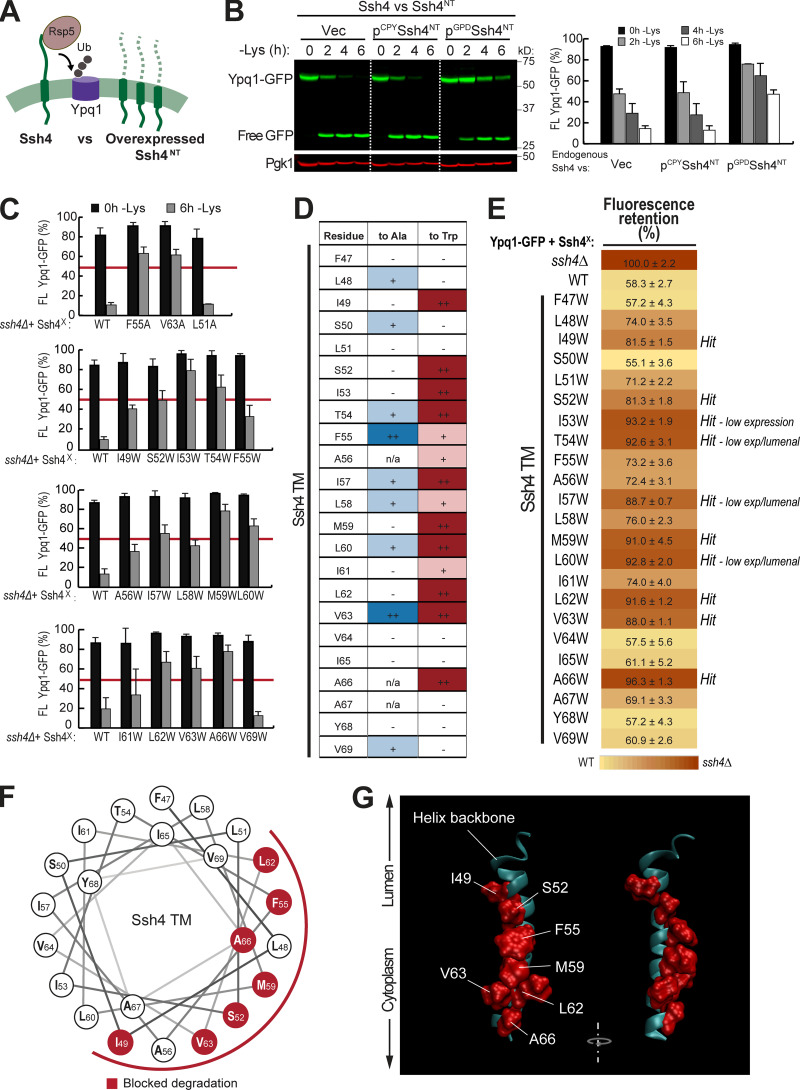
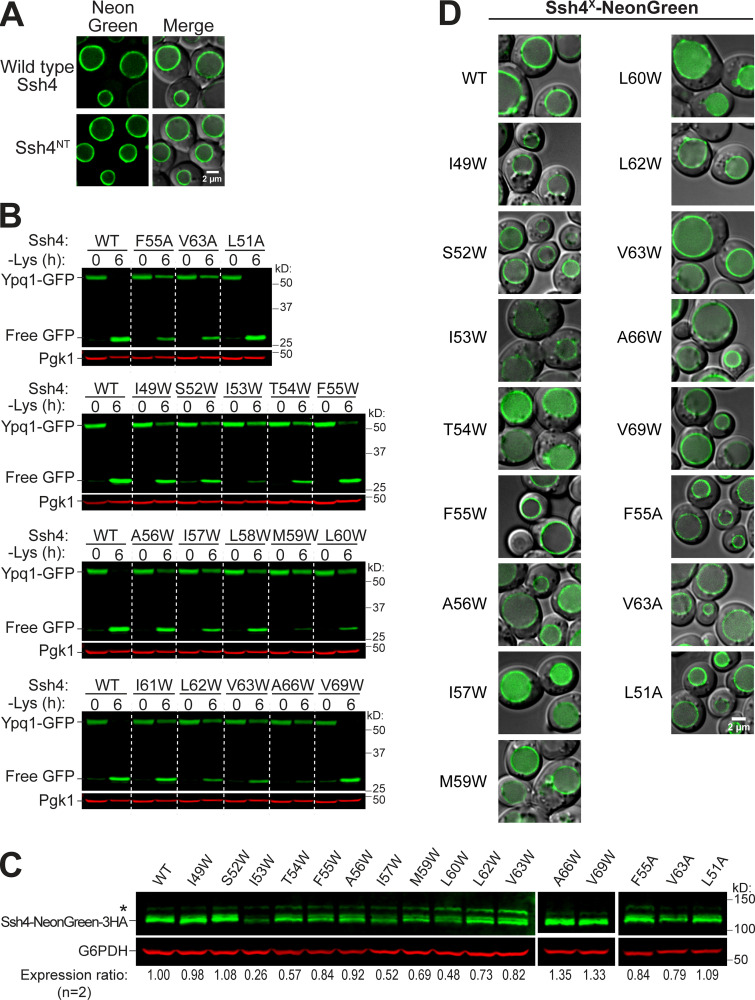
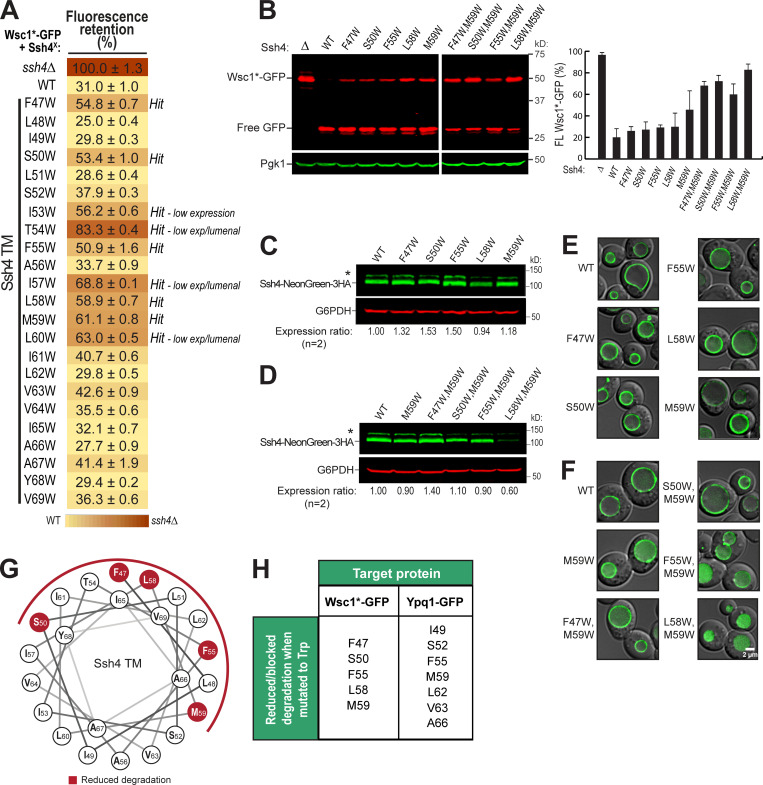
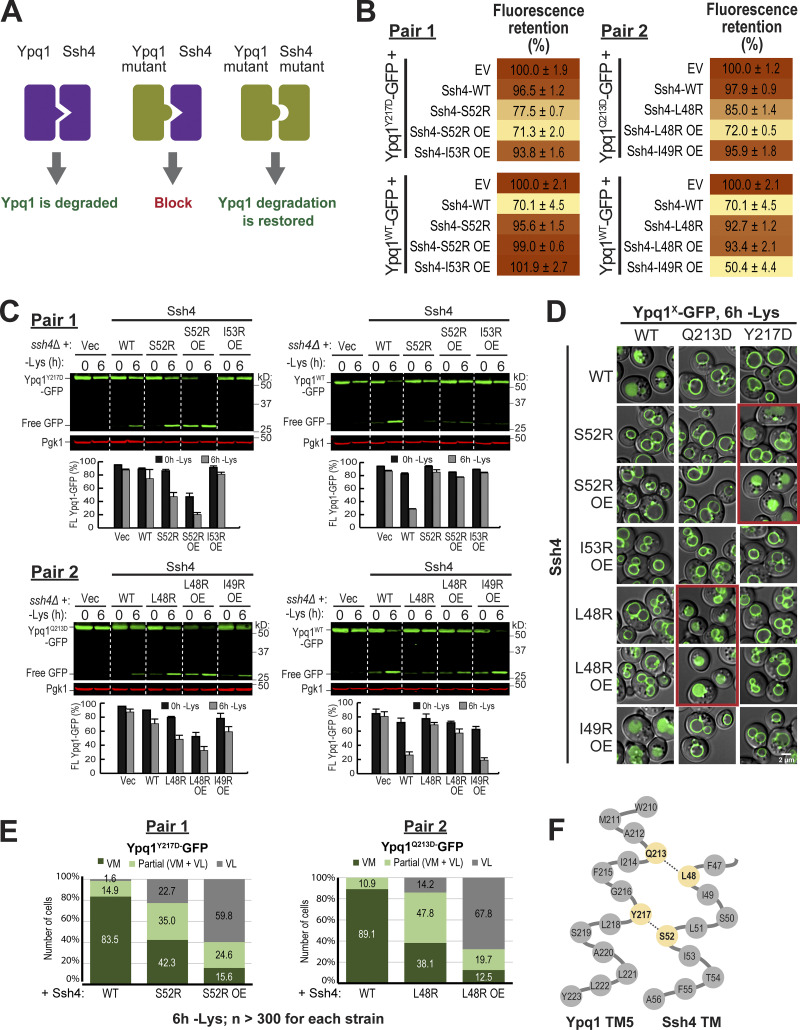
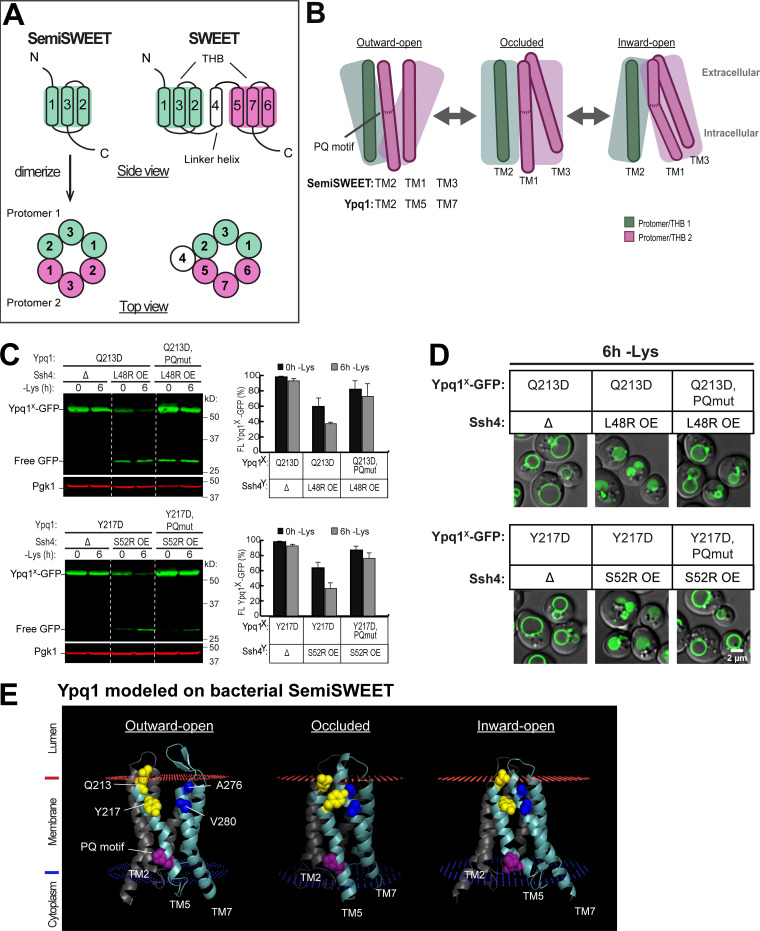
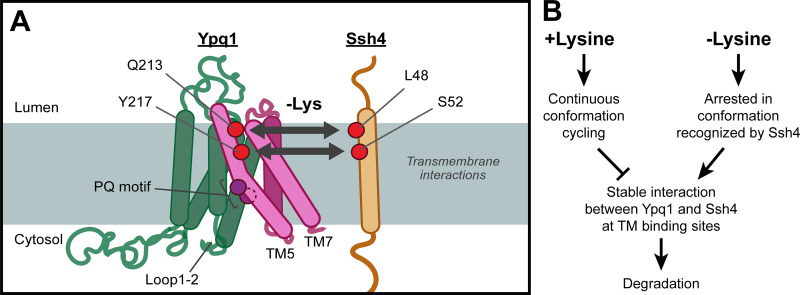
While it is well-known that E3 ubiquitin ligases can selectively ubiquitinate membrane proteins in response to specific environmental cues, the underlying mechanisms for the selectivity are poorly understood. In particular, the role of transmembrane regions, if any, in target recognition remains an open question. Here, we describe how Ssh4, a yeast E3 ligase adaptor, recognizes the PQ-loop lysine transporter Ypq1 only after lysine starvation. We show evidence of an interaction between two transmembrane helices of Ypq1 (TM5 and TM7) and the single transmembrane helix of Ssh4. This interaction is regulated by the conserved PQ motif. Strikingly, recent structural studies of the PQ-loop family have suggested that TM5 and TM7 undergo major conformational changes during substrate transport, implying that transport-associated conformational changes may determine the selectivity. These findings thus provide critical information concerning the regulatory mechanism through which transmembrane domains can be specifically recognized in response to changing environmental conditions.
© 2020 Arines et al.
Figures
Comment in
-
Chaperoning transmembrane helices in the lipid bilayer.J Cell Biol. 2021 Jan 4;220(1):e202012041. doi: 10.1083/jcb.202012041. J Cell Biol. 2021. PMID: 33351098 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
