

The knowns and unknowns of neural adaptations to resistance training
- PMID: 33355714
- PMCID: PMC7892509
- DOI: 10.1007/s00421-020-04567-3
The knowns and unknowns of neural adaptations to resistance training
Abstract
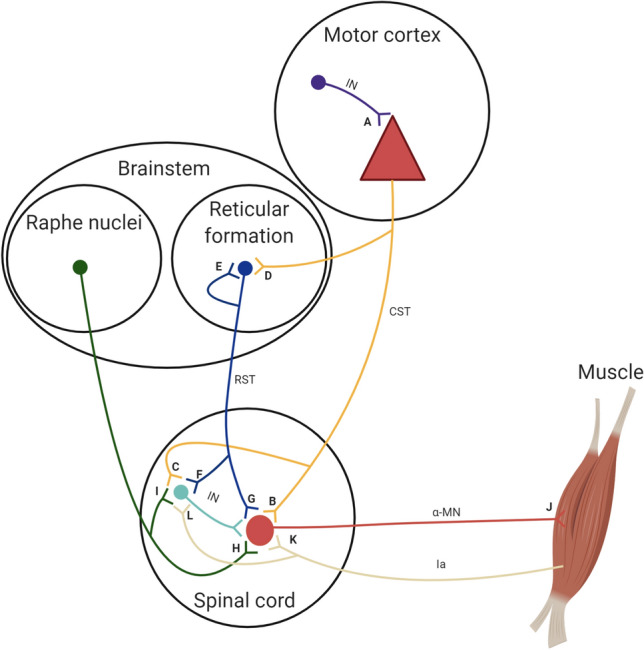
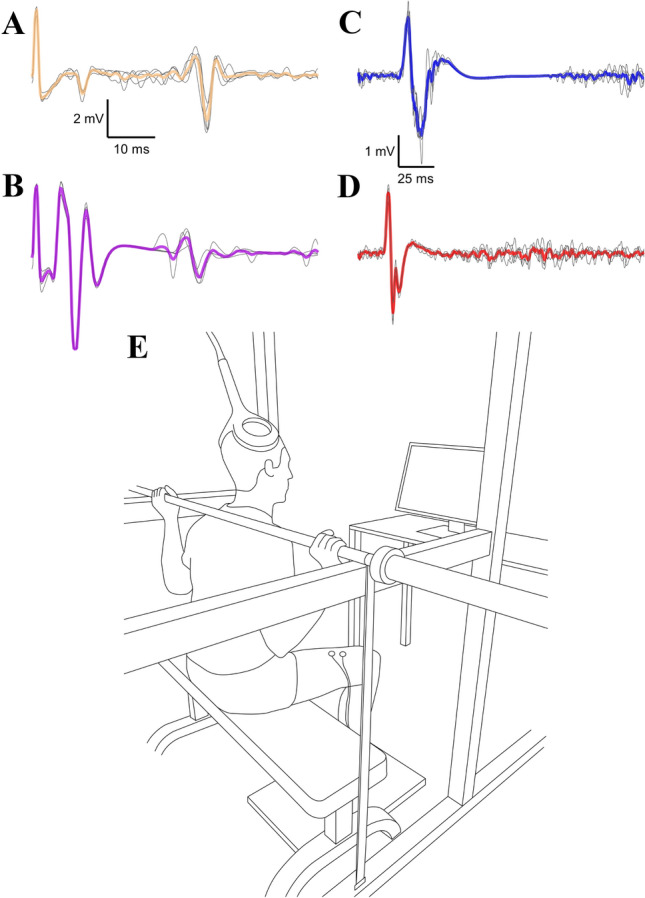
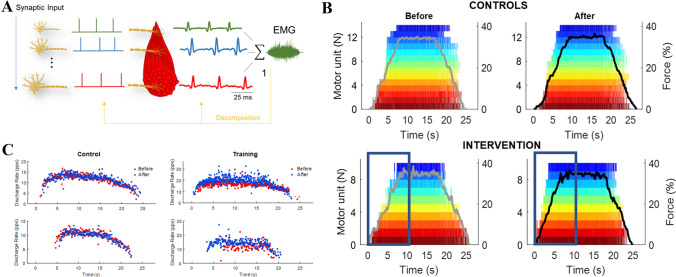
The initial increases in force production with resistance training are thought to be primarily underpinned by neural adaptations. This notion is firmly supported by evidence displaying motor unit adaptations following resistance training; however, the precise locus of neural adaptation remains elusive. The purpose of this review is to clarify and critically discuss the literature concerning the site(s) of putative neural adaptations to short-term resistance training. The proliferation of studies employing non-invasive stimulation techniques to investigate evoked responses have yielded variable results, but generally support the notion that resistance training alters intracortical inhibition. Nevertheless, methodological inconsistencies and the limitations of techniques, e.g. limited relation to behavioural outcomes and the inability to measure volitional muscle activity, preclude firm conclusions. Much of the literature has focused on the corticospinal tract; however, preliminary research in non-human primates suggests reticulospinal tract is a potential substrate for neural adaptations to resistance training, though human data is lacking due to methodological constraints. Recent advances in technology have provided substantial evidence of adaptations within a large motor unit population following resistance training. However, their activity represents the transformation of afferent and efferent inputs, making it challenging to establish the source of adaptation. Whilst much has been learned about the nature of neural adaptations to resistance training, the puzzle remains to be solved. Additional analyses of motoneuron firing during different training regimes or coupling with other methodologies (e.g., electroencephalography) may facilitate the estimation of the site(s) of neural adaptations to resistance training in the future.
Keywords: Descending tracts; High-density surface electromyography; Motor cortex; Motor neuron; Strength; Synaptic input; Transcranial magnetic stimulation.
Conflict of interest statement
The authors declare no conflict of interest, financial or otherwise.
Figures
References
-
- Avela J, Gruber M. Transcranial magnetic stimulation as a tool to study the role of motor cortex in human muscle function. In: Komi P, editor. Neuromuscular aspects of sport performance. Hoboken: Wiley-Blackwell; 2011. pp. 115–134.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
