

Graded titin cleavage progressively reduces tension and uncovers the source of A-band stability in contracting muscle
- PMID: 33357376
- PMCID: PMC7781594
- DOI: 10.7554/eLife.64107
Graded titin cleavage progressively reduces tension and uncovers the source of A-band stability in contracting muscle
Abstract
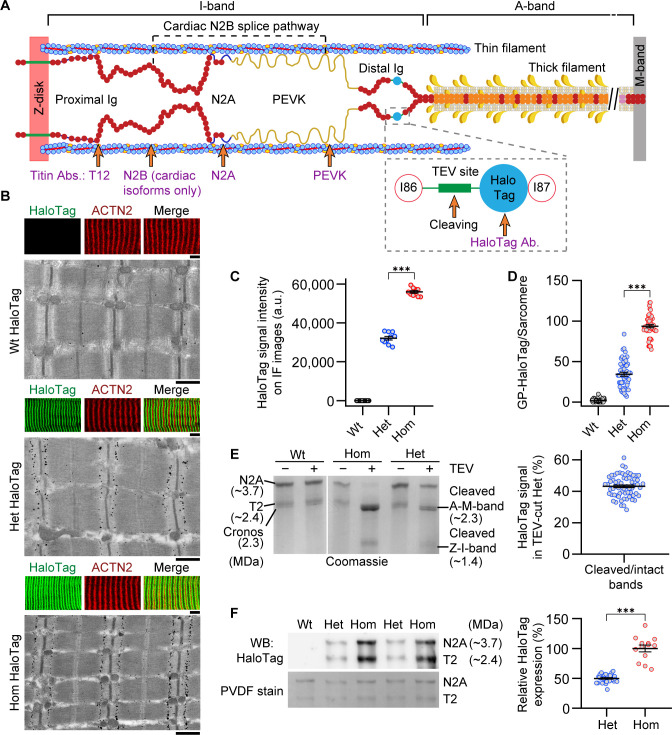
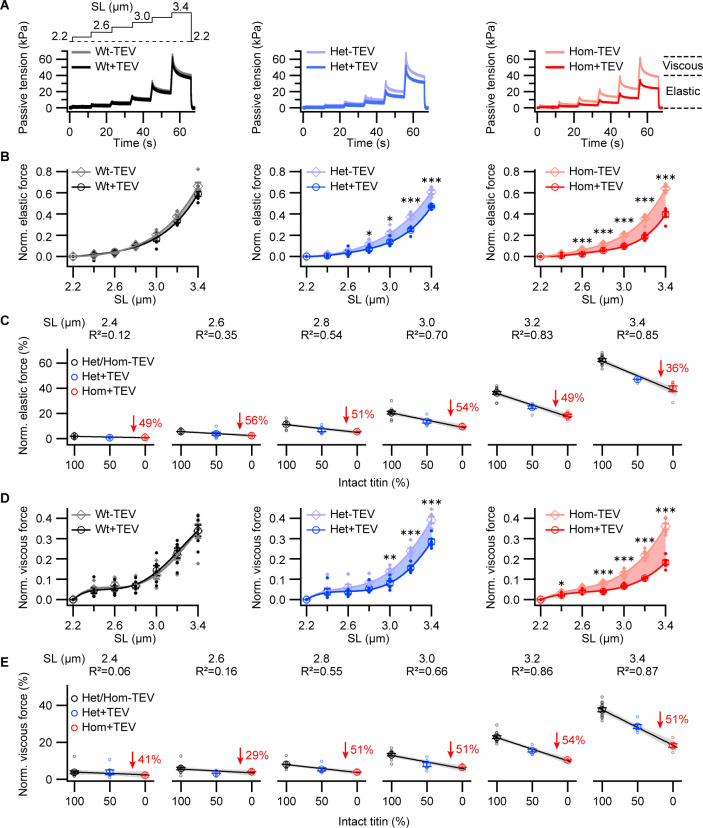
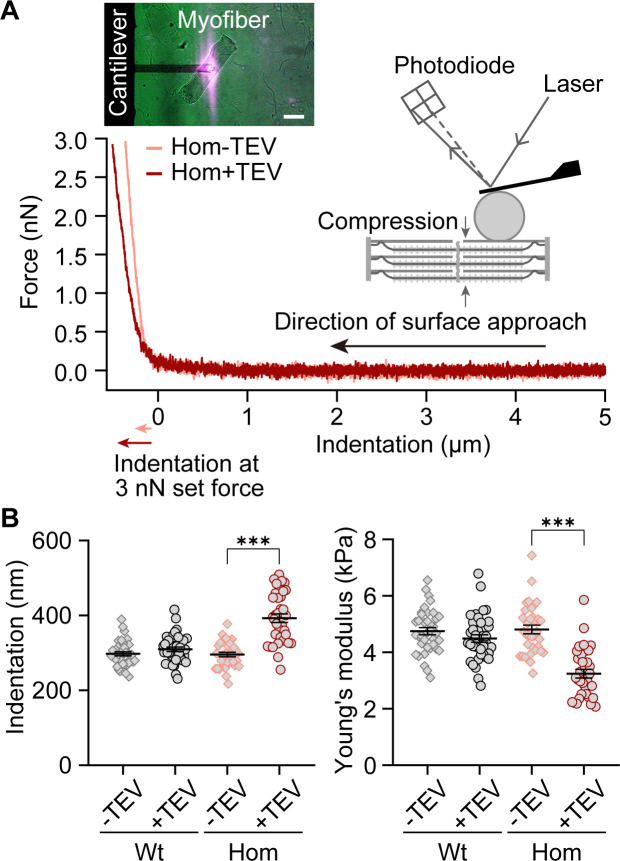
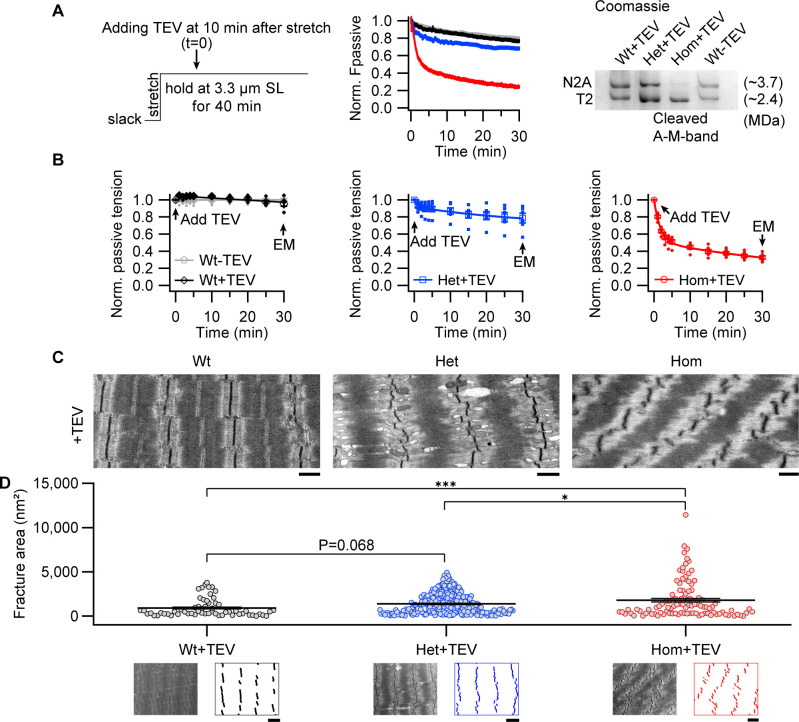
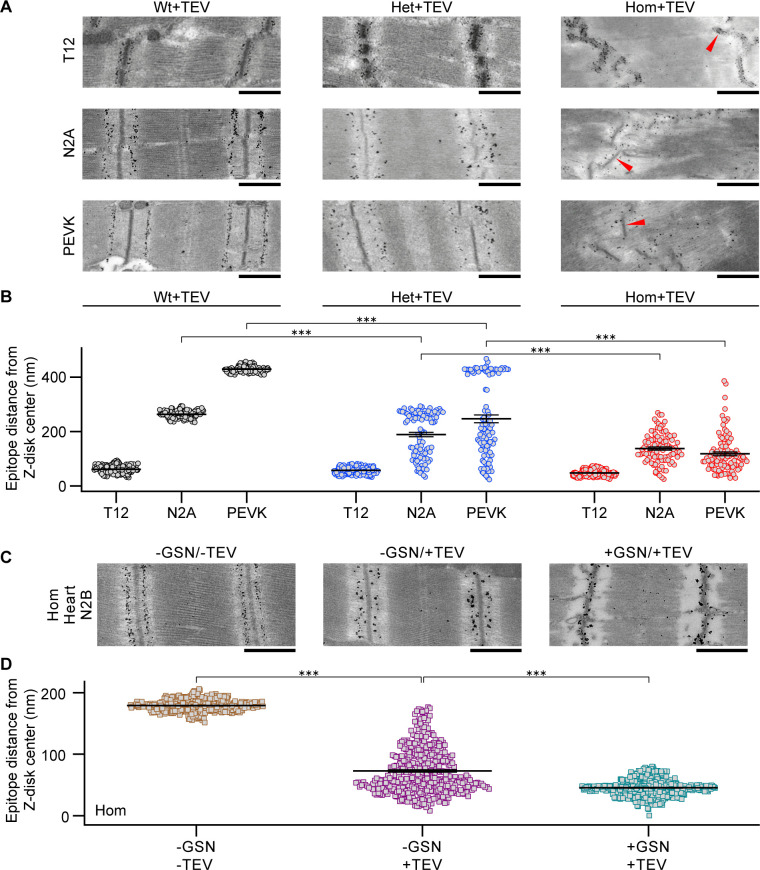
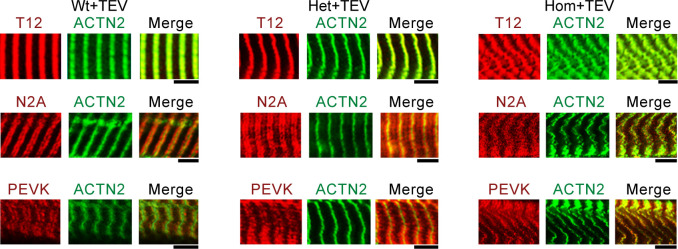
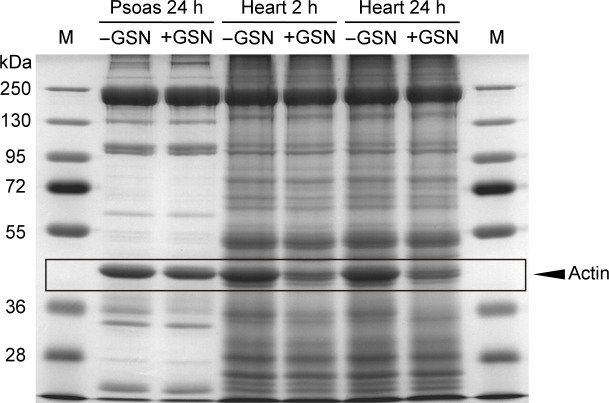
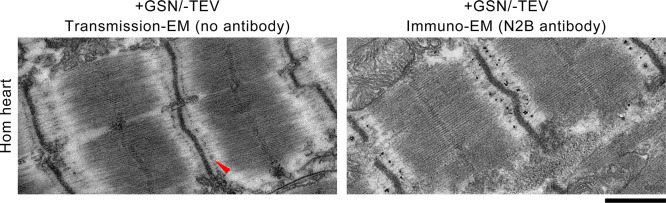
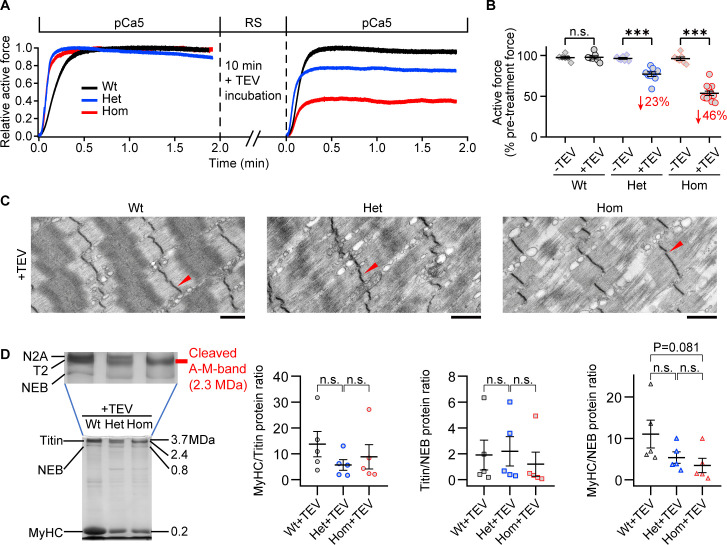
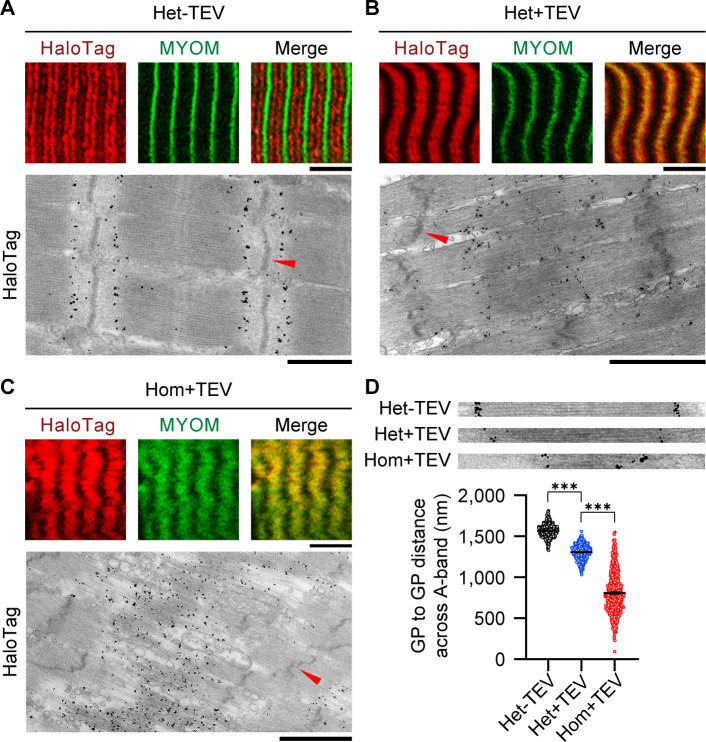
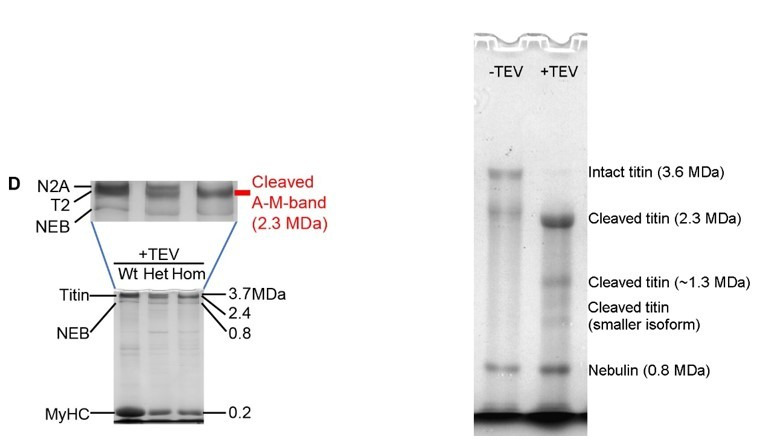
The giant muscle protein titin is a major contributor to passive force; however, its role in active force generation is unresolved. Here, we use a novel titin-cleavage (TC) mouse model that allows specific and rapid cutting of elastic titin to quantify how titin-based forces define myocyte ultrastructure and mechanics. We show that under mechanical strain, as TC doubles from heterozygous to homozygous TC muscles, Z-disks become increasingly out of register while passive and active forces are reduced. Interactions of elastic titin with sarcomeric actin filaments are revealed. Strikingly, when titin-cleaved muscles contract, myosin-containing A-bands become split and adjacent myosin filaments move in opposite directions while also shedding myosins. This establishes intact titin filaments as critical force-transmission networks, buffering the forces observed by myosin filaments during contraction. To perform this function, elastic titin must change stiffness or extensible length, unveiling its fundamental role as an activation-dependent spring in contracting muscle.
Keywords: cell biology; contraction; elasticity; immunocytochemistry; mechanics; mouse; physics of living systems; skeletal muscle fibers; ultrastructure.
© 2020, Li et al.
Conflict of interest statement
YL, AH, AU, DI, JR, FK, JF, WL No competing interests declared
Figures

References
-
- Bang ML, Centner T, Fornoff F, Geach AJ, Gotthardt M, McNabb M, Witt CC, Labeit D, Gregorio CC, Granzier H, Labeit S. The complete gene sequence of titin, expression of an unusual approximately 700-kDa titin isoform, and its interaction with obscurin identify a novel Z-line to I-band linking system. Circulation Research. 2001;89:1065–1072. doi: 10.1161/hh2301.100981. - DOI - PubMed
-
- Brynnel A, Hernandez Y, Kiss B, Lindqvist J, Adler M, Kolb J, van der Pijl R, Gohlke J, Strom J, Smith J, Ottenheijm C, Granzier HL. Downsizing the molecular spring of the giant protein titin reveals that skeletal muscle titin determines passive stiffness and drives longitudinal hypertrophy. eLife. 2018;7:e40532. doi: 10.7554/eLife.40532. - DOI - PMC - PubMed
-
- Buck D, Smith JE, Chung CS, Ono Y, Sorimachi H, Labeit S, Granzier HL. Removal of immunoglobulin-like domains from titin’s spring segment alters titin splicing in mouse skeletal muscle and causes myopathy. Journal of General Physiology. 2014;143:215–230. doi: 10.1085/jgp.201311129. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
