

Cell-Type-Specific Immune Dysregulation in Severely Ill COVID-19 Patients
- PMID: 33357411
- PMCID: PMC7744012
- DOI: 10.1016/j.celrep.2020.108590
Cell-Type-Specific Immune Dysregulation in Severely Ill COVID-19 Patients
Erratum in
-
Cell-type-specific immune dysregulation in severely ill COVID-19 patients.Cell Rep. 2021 Mar 30;34(13):108943. doi: 10.1016/j.celrep.2021.108943. Cell Rep. 2021. PMID: 33789116 Free PMC article. No abstract available.
Abstract
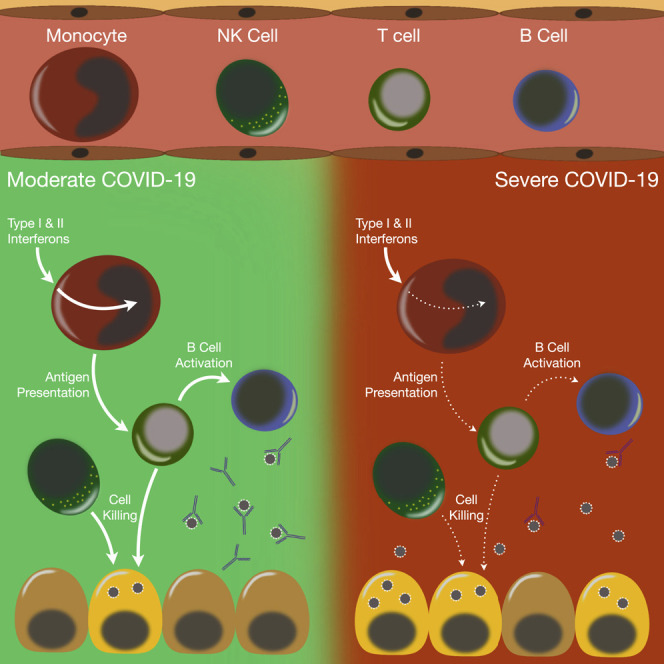
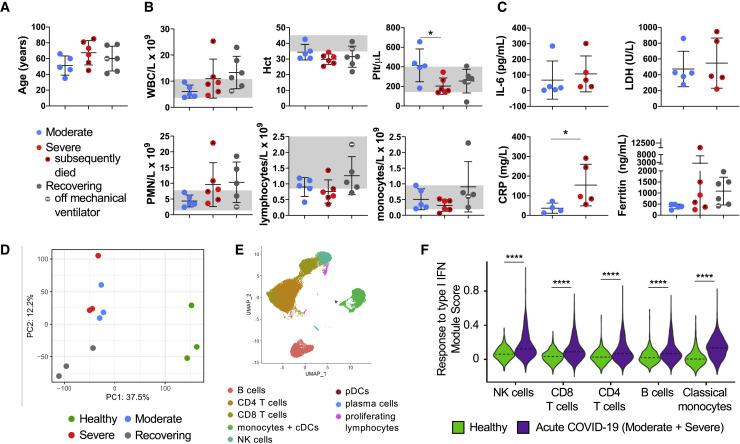
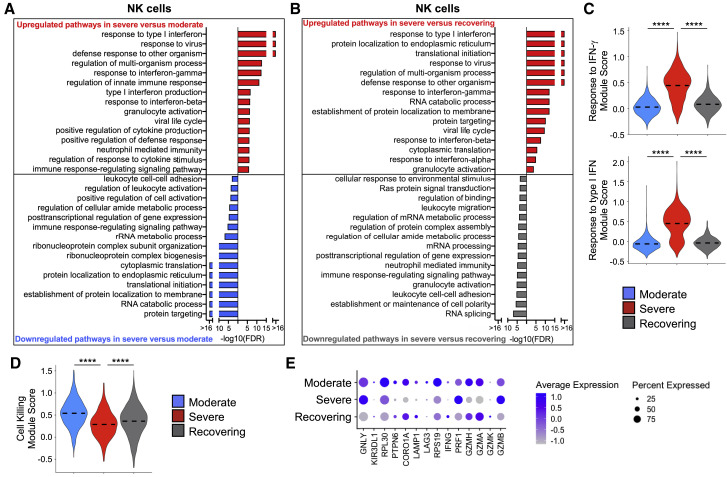
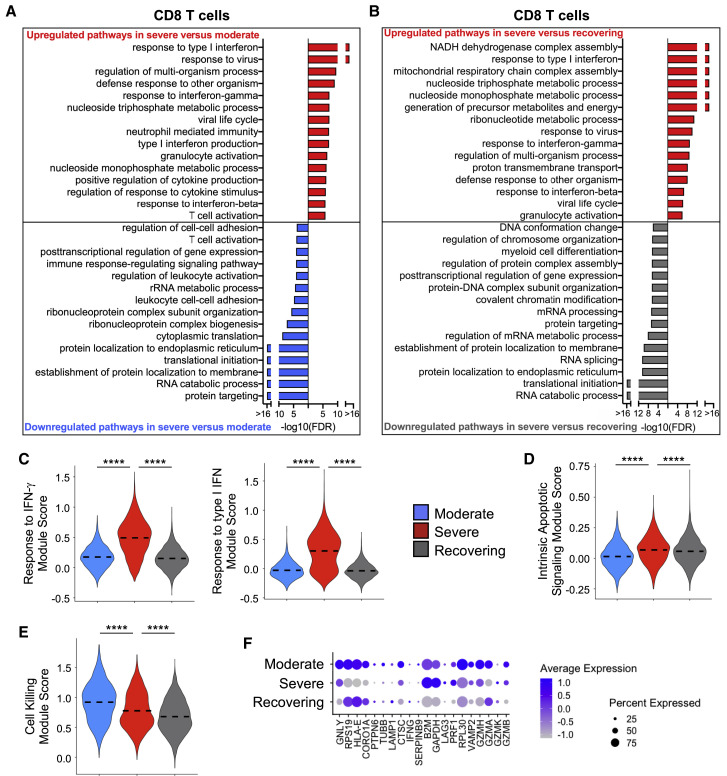
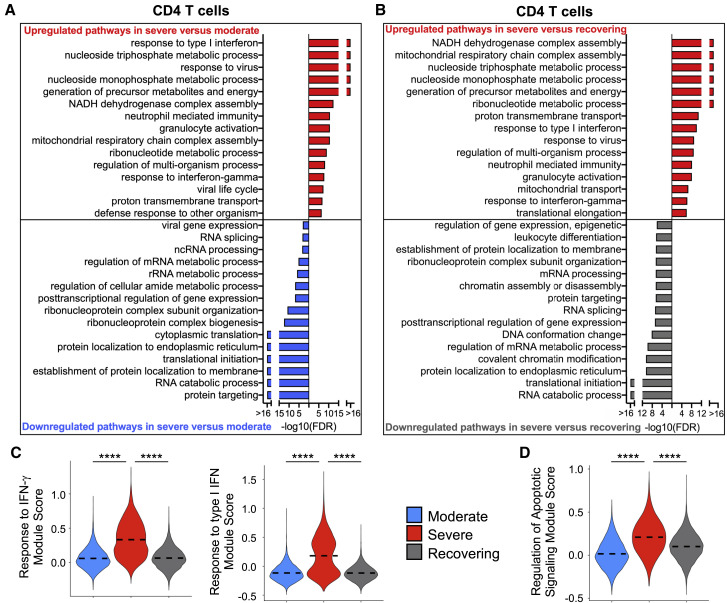
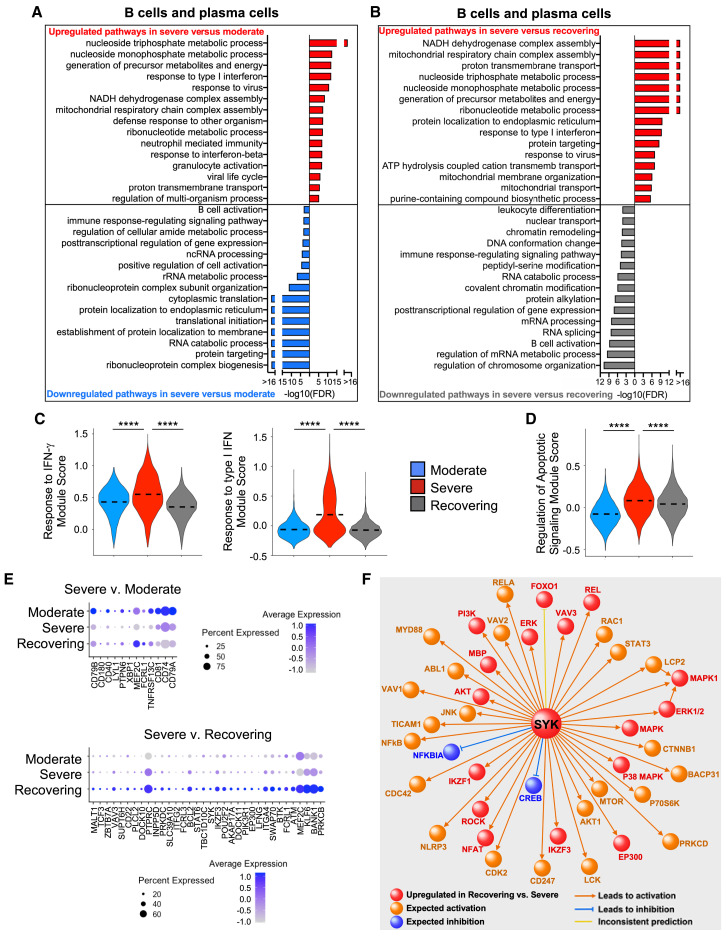
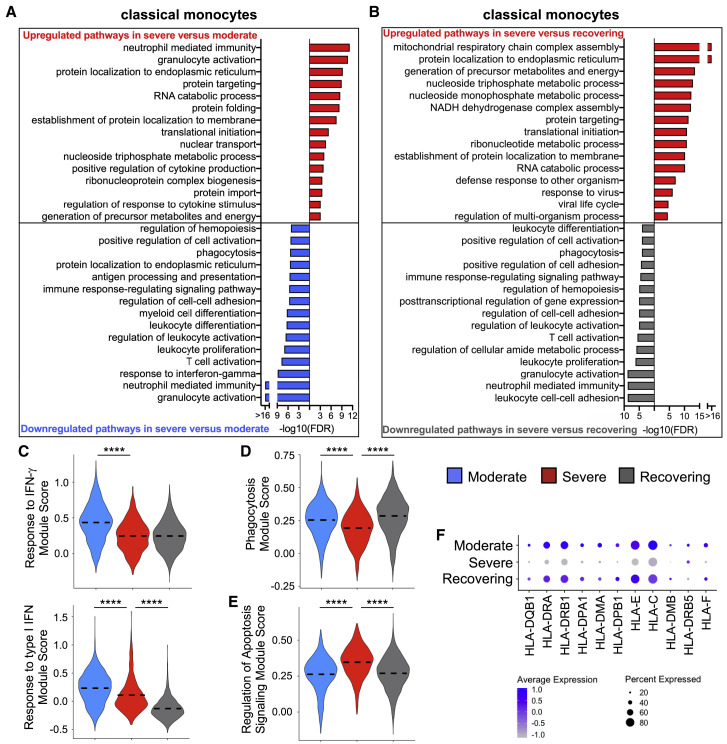
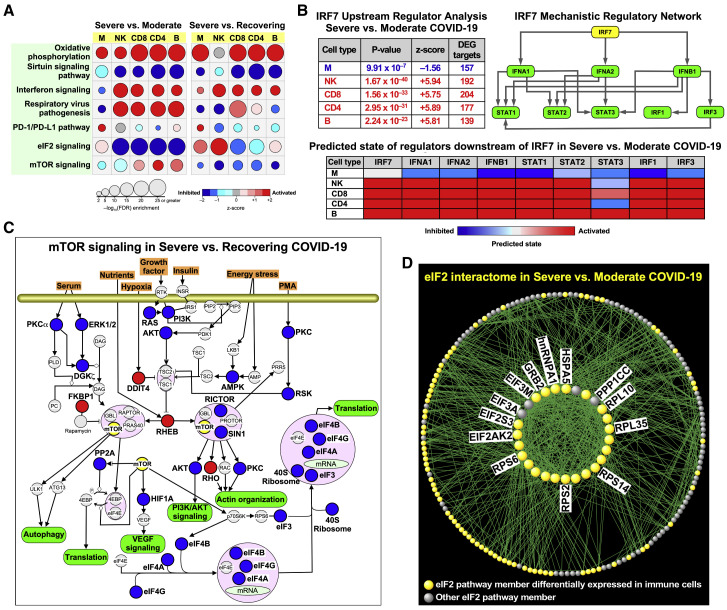
Recent studies have demonstrated immunologic dysfunction in severely ill coronavirus disease 2019 (COVID-19) patients. We use single-cell RNA sequencing (scRNA-seq) to analyze the transcriptome of peripheral blood mononuclear cells (PBMCs) from healthy (n = 3) and COVID-19 patients with moderate disease (n = 5), acute respiratory distress syndrome (ARDS, n = 6), or recovering from ARDS (n = 6). Our data reveal transcriptomic profiles indicative of defective antigen presentation and interferon (IFN) responsiveness in monocytes from ARDS patients, which contrasts with higher responsiveness to IFN signaling in lymphocytes. Furthermore, genes involved in cytotoxic activity are suppressed in both natural killer (NK) and CD8 T lymphocytes, and B cell activation is deficient, which is consistent with delayed viral clearance in severely ill COVID-19 patients. Our study demonstrates that COVID-19 patients with ARDS have a state of immune imbalance in which dysregulation of both innate and adaptive immune responses may be contributing to a more severe disease course.
Keywords: ARDS; COVID-19; SARS-CoV-2; acute respiratory distress syndrome; coronavirus disease 2019; network analysis; severe acute respiratory syndrome coronavirus 2; single-cell RNA sequencing.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Update of
-
Cell type-specific immune dysregulation in severely ill COVID-19 patients.medRxiv [Preprint]. 2020 Jul 24:2020.07.23.20161182. doi: 10.1101/2020.07.23.20161182. medRxiv. 2020. Update in: Cell Rep. 2021 Jan 5;34(1):108590. doi: 10.1016/j.celrep.2020.108590. PMID: 32743611 Free PMC article. Updated. Preprint.
Similar articles
-
Cell type-specific immune dysregulation in severely ill COVID-19 patients.medRxiv [Preprint]. 2020 Jul 24:2020.07.23.20161182. doi: 10.1101/2020.07.23.20161182. medRxiv. 2020. Update in: Cell Rep. 2021 Jan 5;34(1):108590. doi: 10.1016/j.celrep.2020.108590. PMID: 32743611 Free PMC article. Updated. Preprint.
-
Comparative immune profiling of acute respiratory distress syndrome patients with or without SARS-CoV-2 infection.Cell Rep Med. 2021 Jun 15;2(6):100291. doi: 10.1016/j.xcrm.2021.100291. Epub 2021 May 6. Cell Rep Med. 2021. PMID: 33977279 Free PMC article.
-
Lymphocyte Subset Alteration and Monocyte CD4 Expression Reduction in Patients with Severe COVID-19.Viral Immunol. 2021 Jun;34(5):342-351. doi: 10.1089/vim.2020.0166. Epub 2020 Nov 23. Viral Immunol. 2021. PMID: 33264073
-
Immunothrombosis in Acute Respiratory Dysfunction of COVID-19.Front Immunol. 2021 Jun 2;12:651545. doi: 10.3389/fimmu.2021.651545. eCollection 2021. Front Immunol. 2021. PMID: 34149692 Free PMC article.
-
The Role of Macrophages in the Pathogenesis of SARS-CoV-2-Associated Acute Respiratory Distress Syndrome.Front Immunol. 2021 May 10;12:682871. doi: 10.3389/fimmu.2021.682871. eCollection 2021. Front Immunol. 2021. PMID: 34040616 Free PMC article. Review.
Cited by
-
Epigenetic and transcriptional responses in circulating leukocytes are associated with future decompensation during SARS-CoV-2 infection.iScience. 2023 Nov 29;27(1):108288. doi: 10.1016/j.isci.2023.108288. eCollection 2024 Jan 19. iScience. 2023. PMID: 38179063 Free PMC article.
-
Targeting Inflammasome Activation in COVID-19: Delivery of RNA Interference-Based Therapeutic Molecules.Biomedicines. 2021 Dec 3;9(12):1823. doi: 10.3390/biomedicines9121823. Biomedicines. 2021. PMID: 34944639 Free PMC article. Review.
-
Molecular and Clinical Prognostic Biomarkers of COVID-19 Severity and Persistence.Pathogens. 2022 Mar 2;11(3):311. doi: 10.3390/pathogens11030311. Pathogens. 2022. PMID: 35335635 Free PMC article. Review.
-
Differences in SARS-CoV-2 Vaccine Response Dynamics Between Class-I- and Class-II-Specific T-Cell Receptors in Inflammatory Bowel Disease.Front Immunol. 2022 Apr 8;13:880190. doi: 10.3389/fimmu.2022.880190. eCollection 2022. Front Immunol. 2022. PMID: 35464463 Free PMC article.
-
Mapping disease regulatory circuits at cell-type resolution from single-cell multiomics data.Nat Comput Sci. 2023 Jul;3(7):644-657. doi: 10.1038/s43588-023-00476-5. Epub 2023 Jul 25. Nat Comput Sci. 2023. PMID: 37974651 Free PMC article.
References
-
- Cajander S., Tina E., Bäckman A., Magnuson A., Strålin K., Söderquist B., Källman J. Quantitative Real-Time Polymerase Chain Reaction Measurement of HLA-DRA Gene Expression in Whole Blood Is Highly Reproducible and Shows Changes That Reflect Dynamic Shifts in Monocyte Surface HLA-DR Expression during the Course of Sepsis. PLoS ONE. 2016;11:e0154690. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
