

Aging of the immune system and impaired muscle regeneration: A failure of immunomodulation of adult myogenesis
- PMID: 33359378
- PMCID: PMC7855614
- DOI: 10.1016/j.exger.2020.111200
Aging of the immune system and impaired muscle regeneration: A failure of immunomodulation of adult myogenesis
Abstract
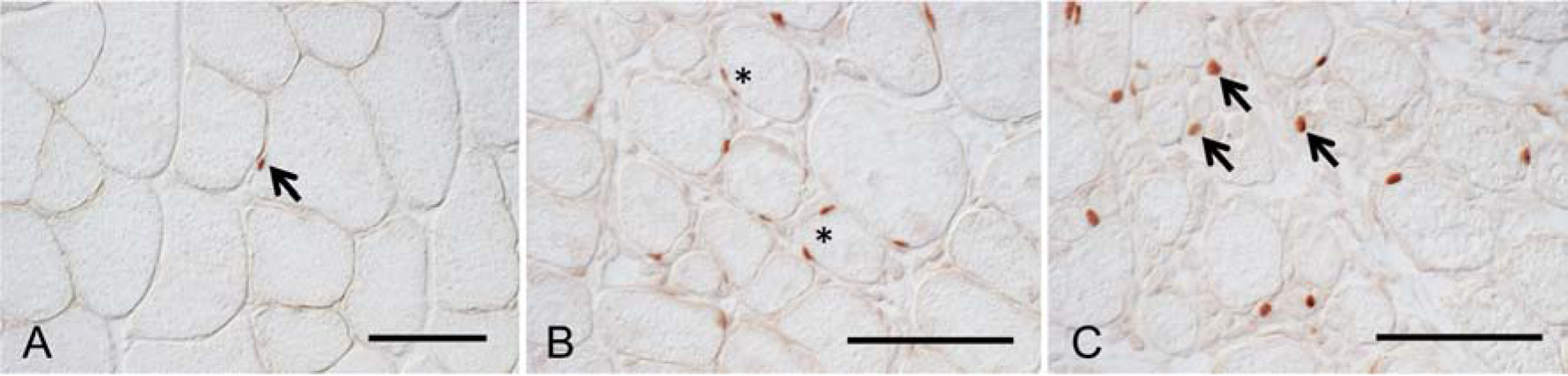
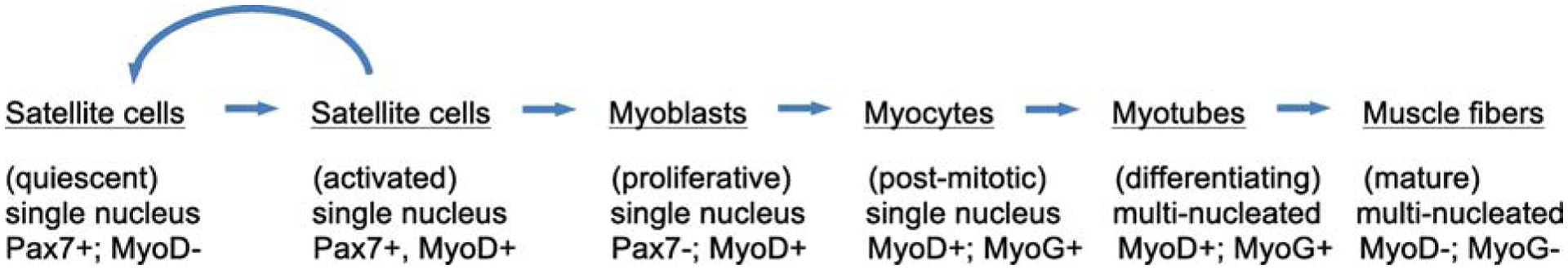
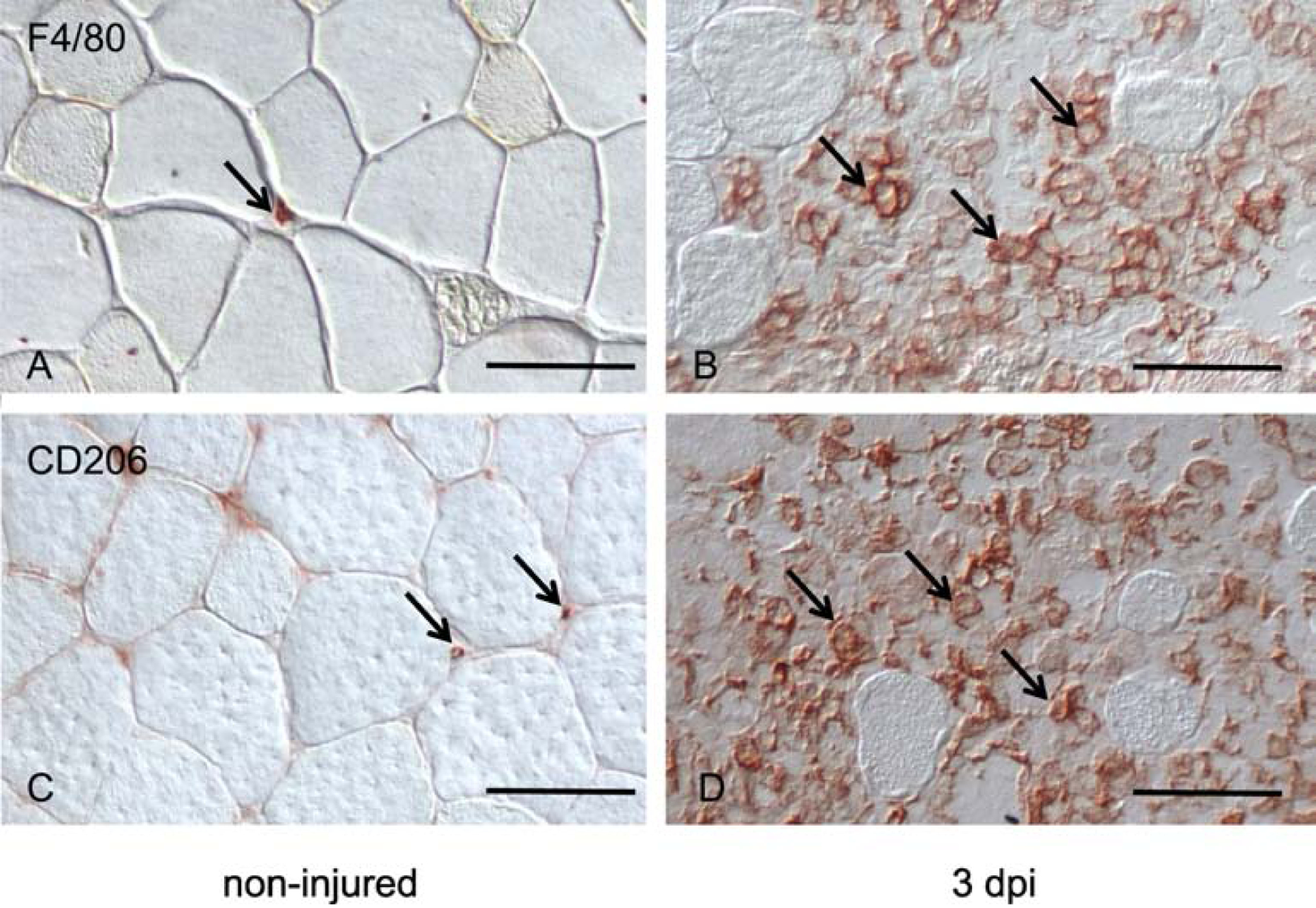
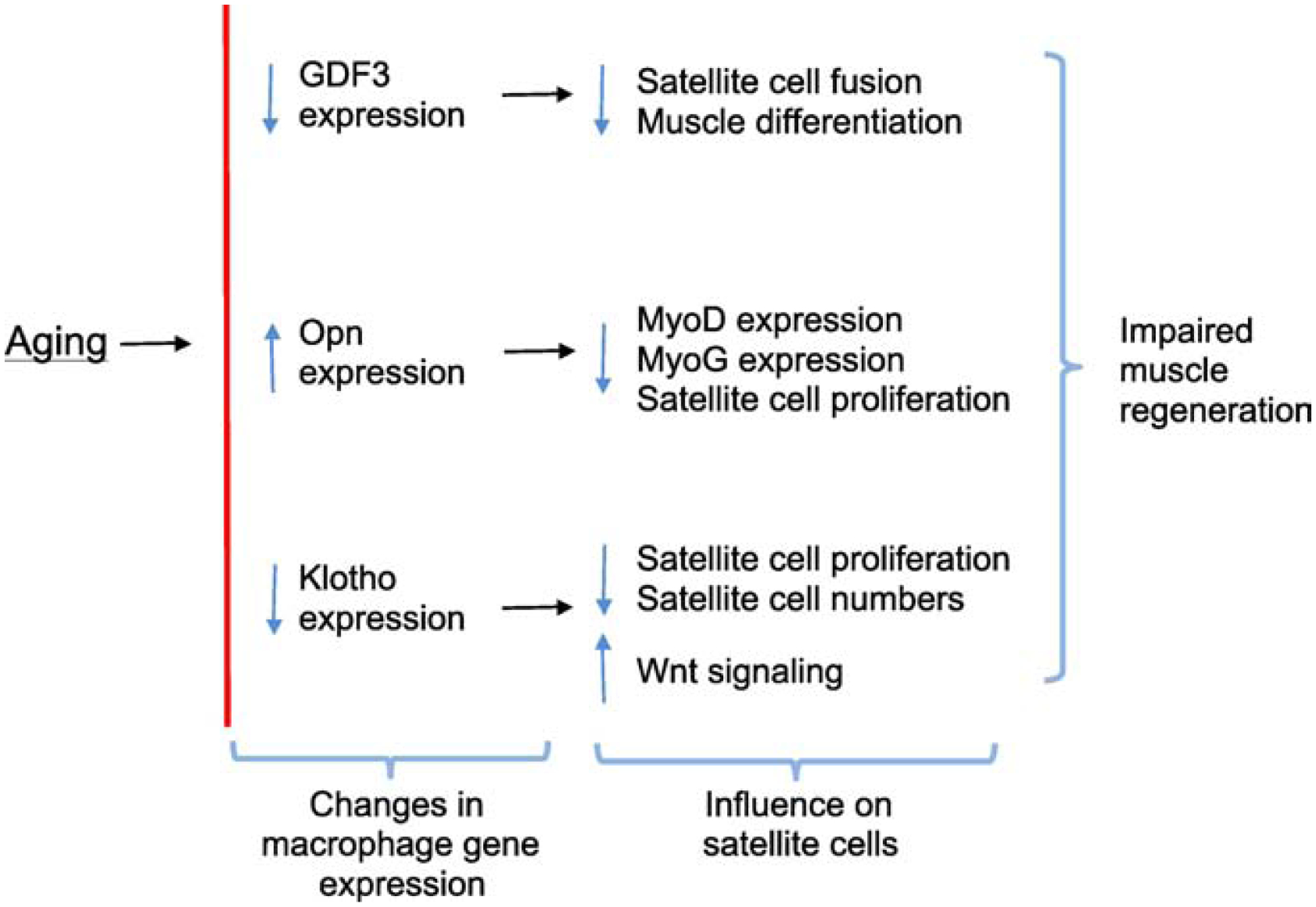
Skeletal muscle regeneration that follows acute injury is strongly influenced by interactions with immune cells that invade and proliferate in the damaged tissue. Discoveries over the past 20 years have identified many of the key mechanisms through which myeloid cells, especially macrophages, regulate muscle regeneration. In addition, lymphoid cells that include CD8+ T-cells and regulatory T-cells also significantly affect the course of muscle regeneration. During aging, the regenerative capacity of skeletal muscle declines, which can contribute to progressive loss of muscle mass and function. Those age-related reductions in muscle regeneration are accompanied by systemic, age-related changes in the immune system, that affect many of the myeloid and lymphoid cell populations that can influence muscle regeneration. In this review, we present recent discoveries that indicate that aging of the immune system contributes to the diminished regenerative capacity of aging muscle. Intrinsic, age-related changes in immune cells modify their expression of factors that affect the function of a population of muscle stem cells, called satellite cells, that are necessary for normal muscle regeneration. For example, age-related reductions in the expression of growth differentiation factor-3 (GDF3) or CXCL10 by macrophages negatively affect adult myogenesis, by disrupting regulatory interactions between macrophages and satellite cells. Those changes contribute to a reduction in the numbers and myogenic capacity of satellite cells in old muscle, which reduces their ability to restore damaged muscle. In addition, aging produces changes in the expression of molecules that regulate the inflammatory response to injured muscle, which also contributes to age-related defects in muscle regeneration. For example, age-related increases in the production of osteopontin by macrophages disrupts the normal inflammatory response to muscle injury, resulting in regenerative defects. These nascent findings represent the beginning of a newly-developing field of investigation into mechanisms through which aging of the immune system affects muscle regeneration.
Keywords: Aging; Muscle inflammation; Muscle injury; Muscle regeneration; Skeletal muscle.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
