

Staphylococcus lugdunensis: a Skin Commensal with Invasive Pathogenic Potential
- PMID: 33361142
- PMCID: PMC7950365
- DOI: 10.1128/CMR.00205-20
Staphylococcus lugdunensis: a Skin Commensal with Invasive Pathogenic Potential
Abstract
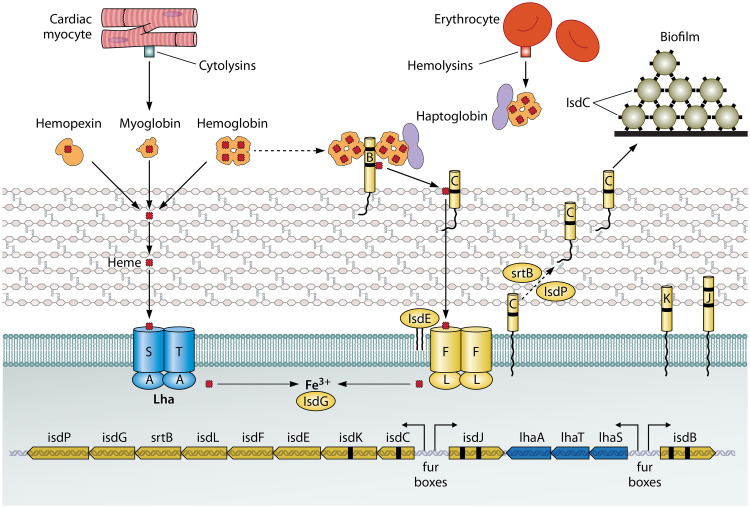
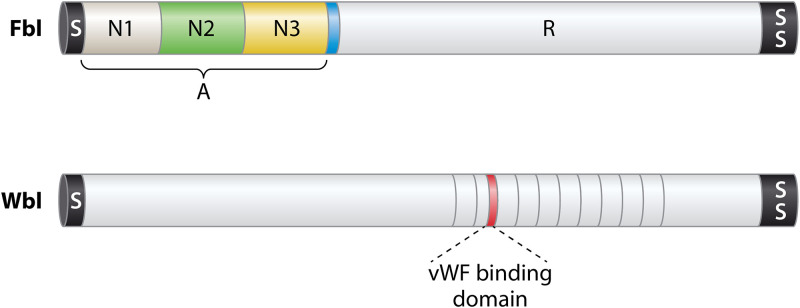
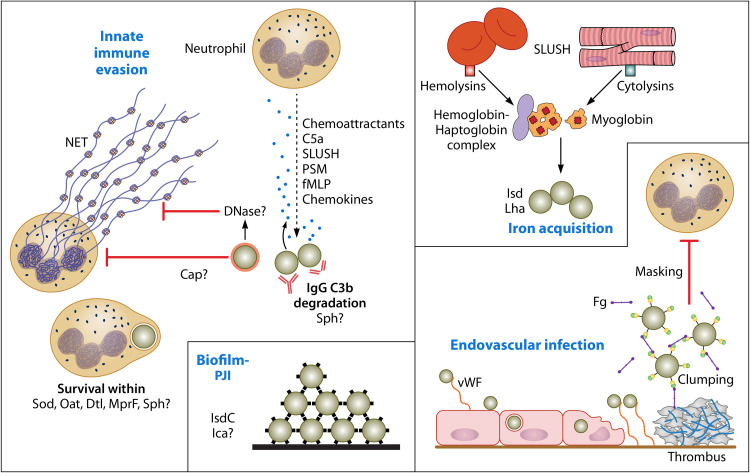
Staphylococcus lugdunensis is a species of coagulase-negative staphylococcus (CoNS) that causes serious infections in humans akin to those of S. aureus It was often misidentified as S. aureus, but this has been rectified by recent routine use of matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) in diagnostic laboratories. It encodes a diverse array of virulence factors for adhesion, cytotoxicity, and innate immune evasion, but these are less diverse than those encoded by S. aureus It expresses an iron-regulated surface determinant (Isd) system combined with a novel energy-coupling factor (ECF) mechanism for extracting heme from hemoproteins. Small cytolytic S. lugdunensis synergistic hemolysins (SLUSH), peptides related to phenol-soluble modulins of S. aureus, act synergistically with β-toxin to lyse erythrocytes. S. lugdunensis expresses a novel peptide antibiotic, lugdunin, that can influence the nasal and skin microbiota. Endovascular infections are initiated by bacterial adherence to fibrinogen promoted by a homologue of Staphylococcus aureus clumping factor A and to von Willebrand factor on damaged endothelium by an uncharacterized mechanism. S. lugdunensis survives within mature phagolysosomes of macrophages without growing and is released only following apoptosis. This differs fundamentally from S. aureus, which actively grows and expresses bicomponent leukotoxins that cause membrane damage and could contribute to survival in the infected host. S. lugdunensis is being investigated as a probiotic to eradicate S. aureus from the nares of carriers. However, this is contraindicated by its innate virulence. Studies to obtain a deeper understanding of S. lugdunensis colonization, virulence, and microbiome interactions are therefore warranted.
Keywords: endocarditis; fibrinogen; heme; invasive infection; macrophage; probiotic; surface proteins; von Willebrand factor.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Fleurette J, Bes M, Brun Y, Freney J, Forey F, Coulet M, Reverdy ME, Etienne J. 1989. Clinical isolates of Staphylococcus lugdunensis and S. schleiferi: bacteriological characteristics and susceptibility to antimicrobial agents. Res Microbiol 140:107–118. doi: 10.1016/0923-2508(89)90044-2. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
