

Alternative DNA Structures In Vivo: Molecular Evidence and Remaining Questions
- PMID: 33361270
- PMCID: PMC8549851
- DOI: 10.1128/MMBR.00110-20
Alternative DNA Structures In Vivo: Molecular Evidence and Remaining Questions
Erratum in
-
Correction for Poggi and Richard, "Alternative DNA Structures In Vivo: Molecular Evidence and Remaining Questions".Microbiol Mol Biol Rev. 2021 Mar 3;85(2):e00025-21. doi: 10.1128/MMBR.00025-21. Print 2021 May 19. Microbiol Mol Biol Rev. 2021. PMID: 33658325 Free PMC article. No abstract available.
Abstract
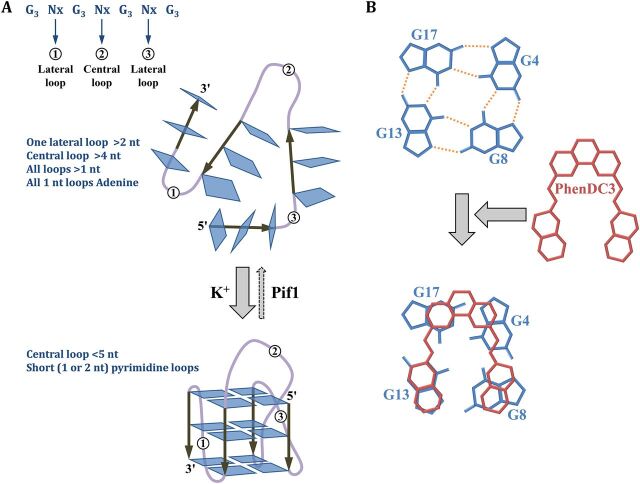
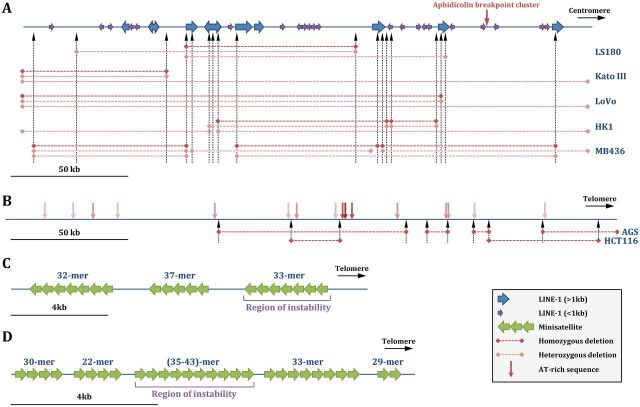
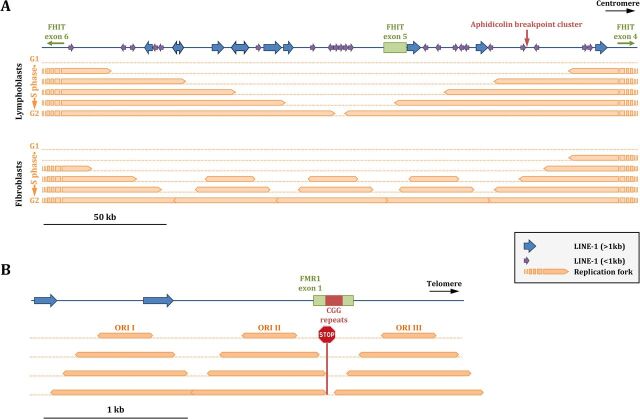
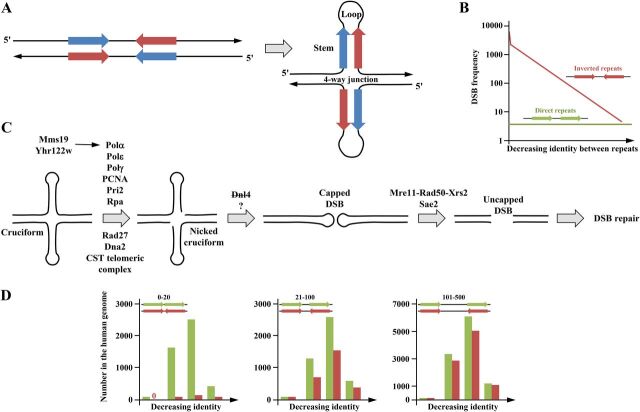
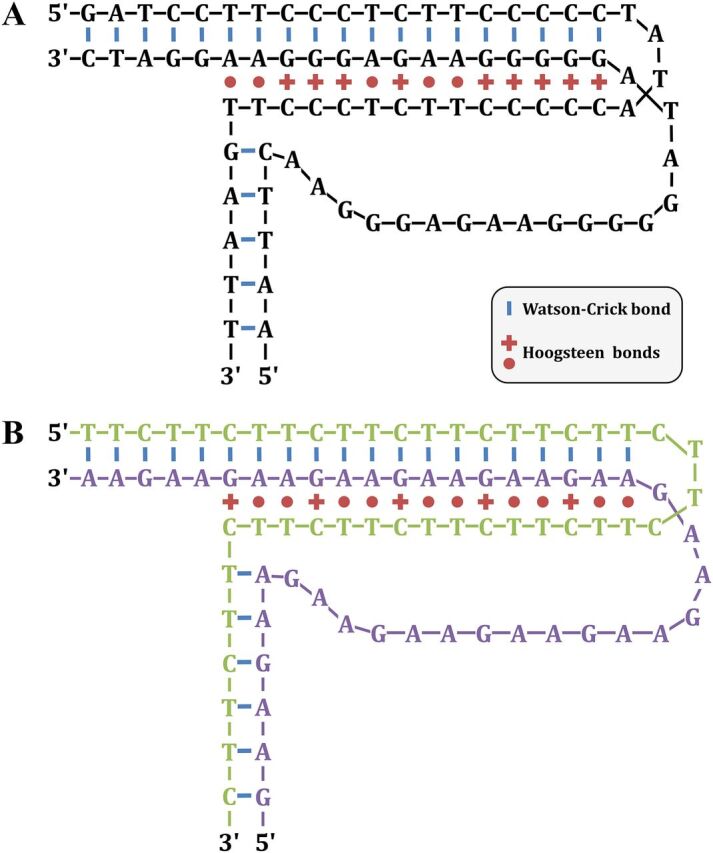
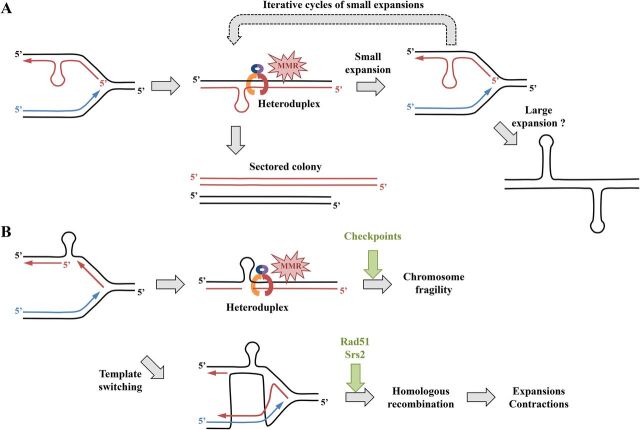
Duplex DNA naturally folds into a right-handed double helix in physiological conditions. Some sequences of unusual base composition may nevertheless form alternative structures, as was shown for many repeated sequences in vitro However, evidence for the formation of noncanonical structures in living cells is difficult to gather. It mainly relies on genetic assays demonstrating their function in vivo or through genetic instability reflecting particular properties of such structures. Efforts were made to reveal their existence directly in a living cell, mainly by generating antibodies specific to secondary structures or using chemical ligands selected for their affinity to these structures. Among secondary structure-forming DNAs are G-quadruplexes, human fragile sites containing minisatellites, AT-rich regions, inverted repeats able to form cruciform structures, hairpin-forming CAG/CTG triplet repeats, and triple helices formed by homopurine-homopyrimidine GAA/TTC trinucleotide repeats. Many of these alternative structures are involved in human pathologies, such as neurological or developmental disorders, as in the case of trinucleotide repeats, or cancers triggered by translocations linked to fragile sites. This review will discuss and highlight evidence supporting the formation of alternative DNA structures in vivo and will emphasize the role of the mismatch repair machinery in binding mispaired DNA duplexes, triggering genetic instability.
Keywords: DNA hairpin; G quadruplex; cruciform; fragile sites; mismatch repair; palindromes; secondary structures; trinucleotide repeats.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Chargaff E, Magasanik B, Vischer E, Green C, Doniger R, Elson D. 1950. Nucleotide composition of pentose nucleic acids from yeast and mammalian tissues. J Biol Chem 186:51–67. - PubMed
-
- Hoogsteen K. 1963. The crystal and molecular structure of a hydrogen-bonded complex between 1-methylthymine and 9-methyladenine. Acta Cryst 16:907–916. doi: 10.1107/S0365110X63002437. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
