

Hormopriming to Mitigate Abiotic Stress Effects: A Case Study of N 9-Substituted Cytokinin Derivatives With a Fluorinated Carbohydrate Moiety
- PMID: 33362831
- PMCID: PMC7758400
- DOI: 10.3389/fpls.2020.599228
Hormopriming to Mitigate Abiotic Stress Effects: A Case Study of N 9-Substituted Cytokinin Derivatives With a Fluorinated Carbohydrate Moiety
Abstract
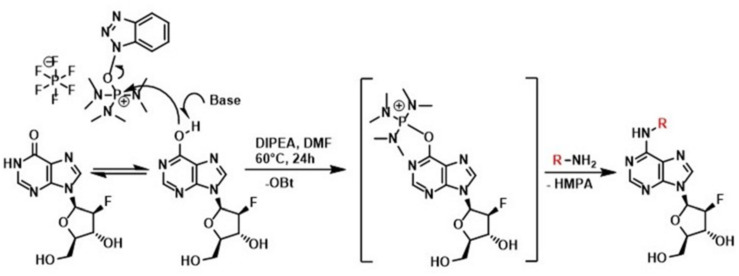
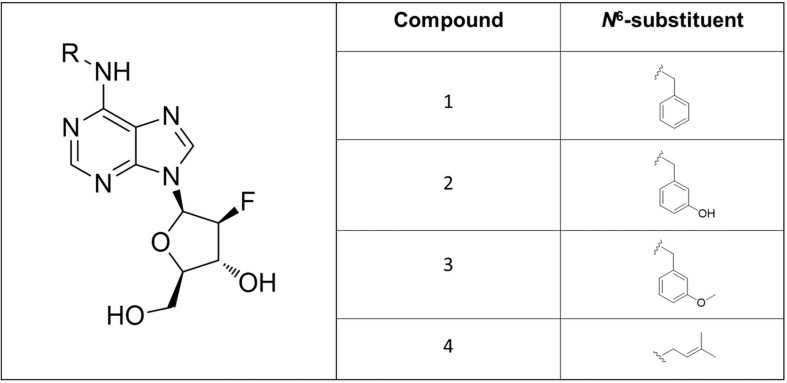
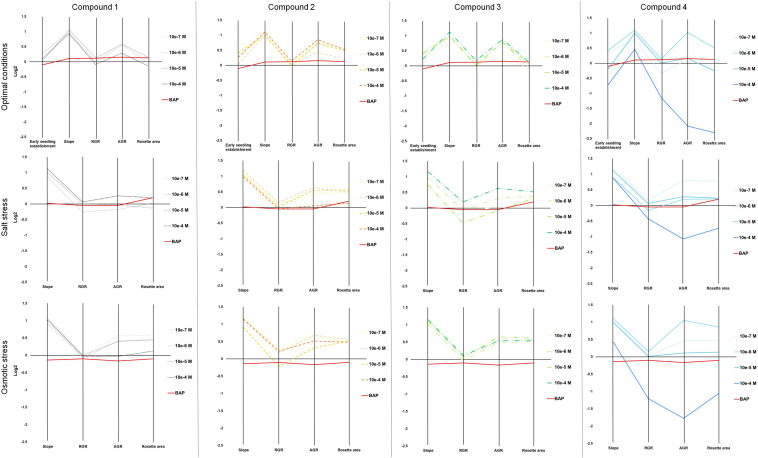
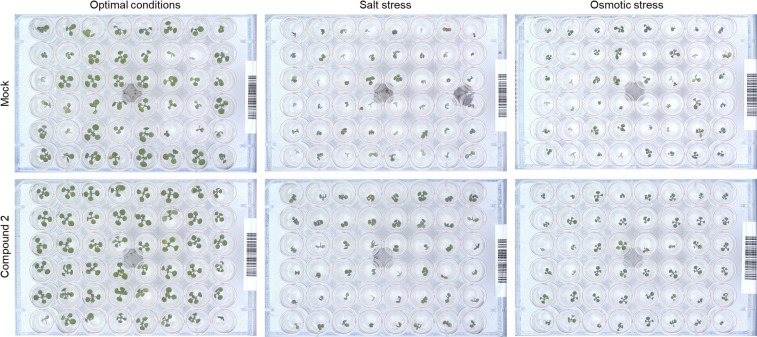
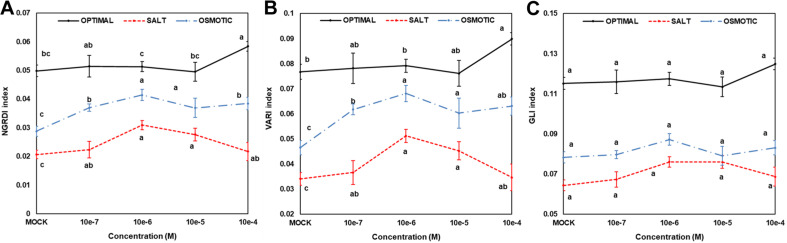
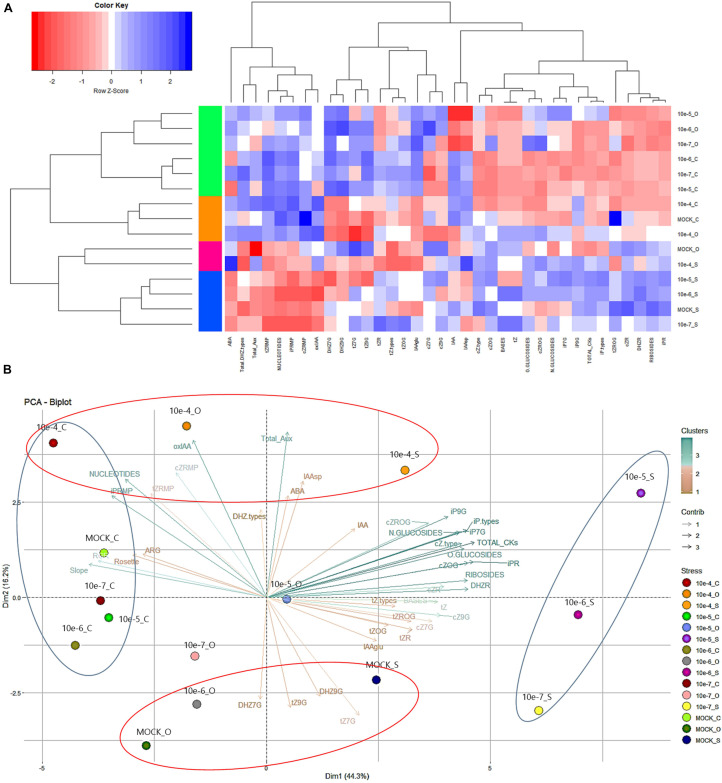
Drought and salinity reduce seed germination, seedling emergence, and early seedling establishment, affect plant metabolism, and hence, reduce crop yield. Development of technologies that can increase plant tolerance of these challenging growth conditions is a major current interest among plant scientists and breeders. Seed priming has become established as one of the practical approaches that can alleviate the negative impact of many environmental stresses and improve the germination and overall performance of crops. Hormopriming using different plant growth regulators has been widely demonstrated as effective, but information about using cytokinins (CKs) as priming agents is limited to only a few studies using kinetin or 6-benzylaminopurine (BAP). Moreover, the mode of action of these compounds in improving seed and plant fitness through priming has not yet been studied. For many years, BAP has been one of the CKs most commonly applied exogenously to plants to delay senescence and reduce the impact of stress. However, rapid endogenous N 9-glucosylation of BAP can result in negative effects. This can be suppressed by hydroxylation of the benzyl ring or by appropriate N 9 purine substitution. Replacement of the 2' or 3' hydroxyl groups of a nucleoside with a fluorine atom has shown promising results in drug research and biochemistry as a means of enhancing biological activity and increasing chemical or metabolic stability. Here, we show that the application of this chemical modification in four new N 9-substituted CK derivatives with a fluorinated carbohydrate moiety improved the antisenescence properties of CKs. Besides, detailed phenotypical analysis of the growth and development of Arabidopsis plants primed with the new CK analogs over a broad concentration range and under various environmental conditions revealed that they improve growth regulation and antistress activity. Seed priming with, for example, 6-(3-hydroxybenzylamino)-2'-deoxy-2'-fluoro-9-(β)-D-arabinofuranosylpurine promoted plant growth under control conditions and alleviated the negative effects of the salt and osmotic stress. The mode of action of this hormopriming and its effect on plant metabolism were further analyzed through quantification of the endogenous levels of phytohormones such as CKs, auxins and abscisic acid, and the results are discussed.
Keywords: Arabidopsis; abiotic stress; antisenescence; cytokinin analogs; hormopriming; plant biostimulant characterization index.
Copyright © 2020 Bryksová, Hybenová, Hernándiz, Novák, Pěnčík, Spíchal, De Diego and Doležal.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Bagheri A., Bagherifard A., Saborifard H., Ahmadi M., Safarpoor M. (2014). Effects drought, cytokinins and GA3 on seedling growth of Basil (Ocimum basilicum). Int. J. Adv. Biol. Biomed. Res. 2 489–493. 10.1017/CBO9781107415324.004 - DOI
-
- Bairu M. W., Jain N., Stirk W. A., Doležal K., Van Staden J. (2009). Solving the problem of shoot-tip necrosis in Harpagophytum procumbens by changing the cytokinin types, calcium and boron concentrations in the medium. South Afr. J. Bot. 75 122–127. 10.1016/j.sajb.2008.08.006 - DOI
-
- Chen K., Arora R. (2013). Priming memory invokes seed stress-tolerance. Environ. Exp. Bot. 94 33–45. 10.1016/j.envexpbot.2012.03.005 - DOI
LinkOut - more resources
Full Text Sources
Research Materials