

Reaction Wood Anatomical Traits and Hormonal Profiles in Poplar Bent Stem and Root
- PMID: 33363556
- PMCID: PMC7754185
- DOI: 10.3389/fpls.2020.590985
Reaction Wood Anatomical Traits and Hormonal Profiles in Poplar Bent Stem and Root
Abstract
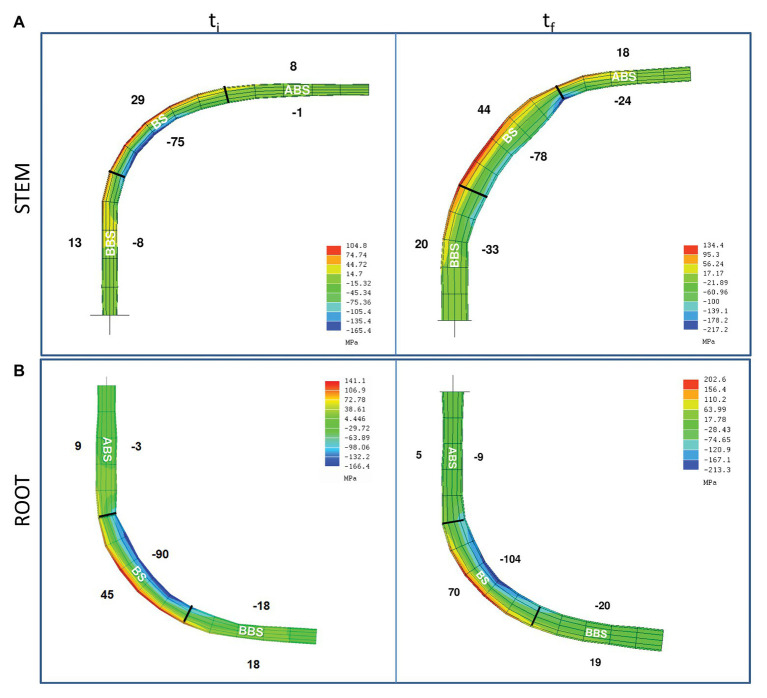
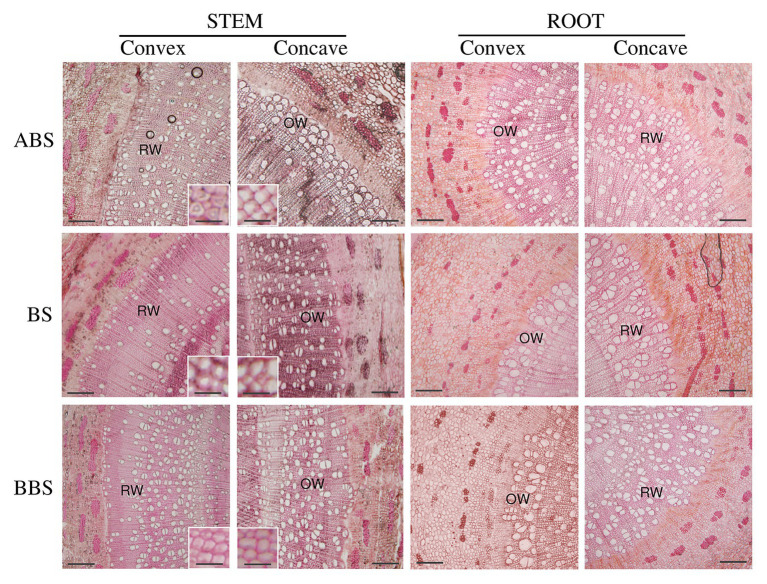
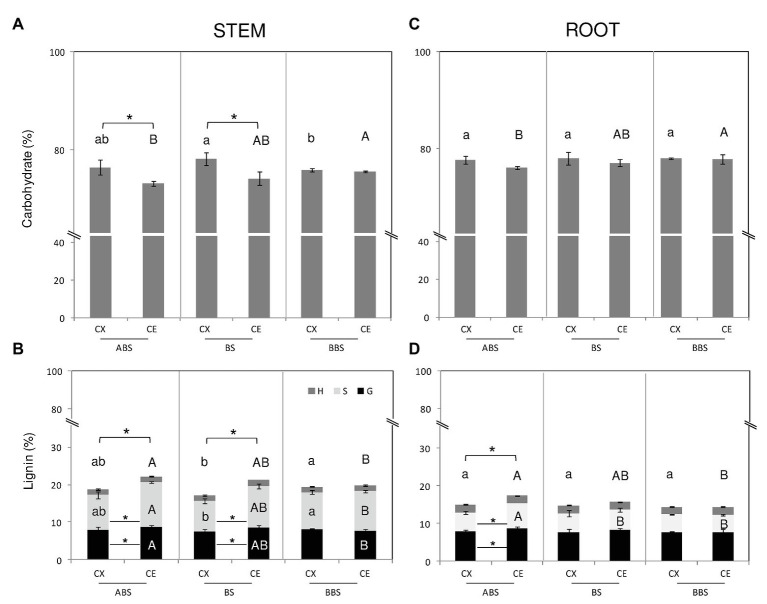
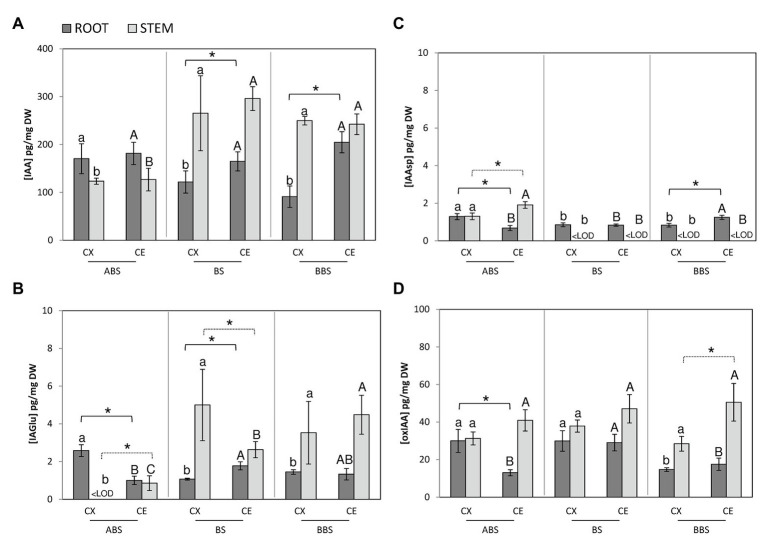
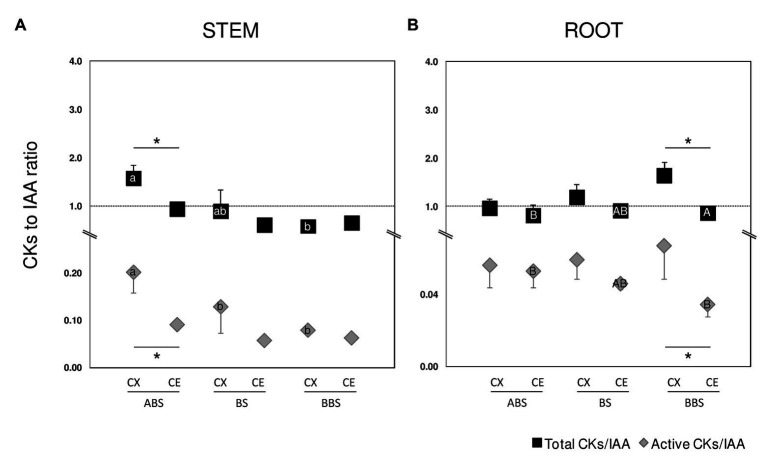
Reaction wood (RW) formation is an innate physiological response of woody plants to counteract mechanical constraints in nature, reinforce structure and redirect growth toward the vertical direction. Differences and/or similarities between stem and root response to mechanical constraints remain almost unknown especially in relation to phytohormones distribution and RW characteristics. Thus, Populus nigra stem and root subjected to static non-destructive mid-term bending treatment were analyzed. The distribution of tension and compression forces was firstly modeled along the main bent stem and root axis; then, anatomical features, chemical composition, and a complete auxin and cytokinin metabolite profiles of the stretched convex and compressed concave side of three different bent stem and root sectors were analyzed. The results showed that in bent stems RW was produced on the upper stretched convex side whereas in bent roots it was produced on the lower compressed concave side. Anatomical features and chemical analysis showed that bent stem RW was characterized by a low number of vessel, poor lignification, and high carbohydrate, and thus gelatinous layer in fiber cell wall. Conversely, in bent root, RW was characterized by high vessel number and area, without any significant variation in carbohydrate and lignin content. An antagonistic interaction of auxins and different cytokinin forms/conjugates seems to regulate critical aspects of RW formation/development in stem and root to facilitate upward/downward organ bending. The observed differences between the response stem and root to bending highlight how hormonal signaling is highly organ-dependent.
Keywords: UHPLC-MS/MS; auxins; bending stress; cytokinins; metabolite profiling.
Copyright © 2020 De Zio, Montagnoli, Karady, Terzaghi, Sferra, Antoniadi, Scippa, Ljung, Chiatante and Trupiano.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Antonova G. F., Varaksina T. N., Zheleznichenko T. V., Stasova V. V. (2014). Lignin deposition during earlywood and latewood formation in scots pine stems. Wood Sci. Technol. 48, 919–936. 10.1007/s00226-014-0650-3 - DOI
-
- Begum S., Nakaba S., Bayramzadeh V., Oribee Y., Kubo T., Funada R. (2008). Temperature responses of cambial reactivation and xylem differentiation in hybrid poplar (Populus sieboldii × P. grandidentata) under natural conditions. Tree Physiol. 28, 1813–1819. 10.1093/treephys/28.12.1813, PMID: - DOI - PubMed
LinkOut - more resources
Full Text Sources