

Molecular Basis for Sesterterpene Diversity Produced by Plant Terpene Synthases
- PMID: 33367256
- PMCID: PMC7747971
- DOI: 10.1016/j.xplc.2020.100051
Molecular Basis for Sesterterpene Diversity Produced by Plant Terpene Synthases
Abstract
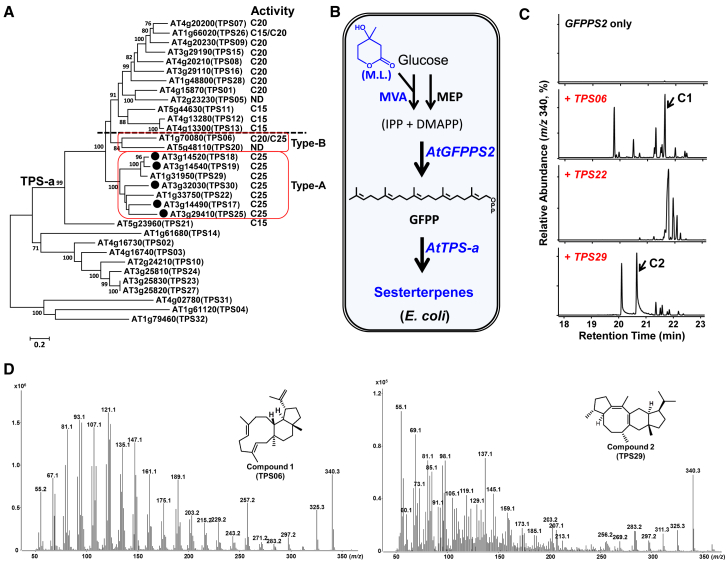
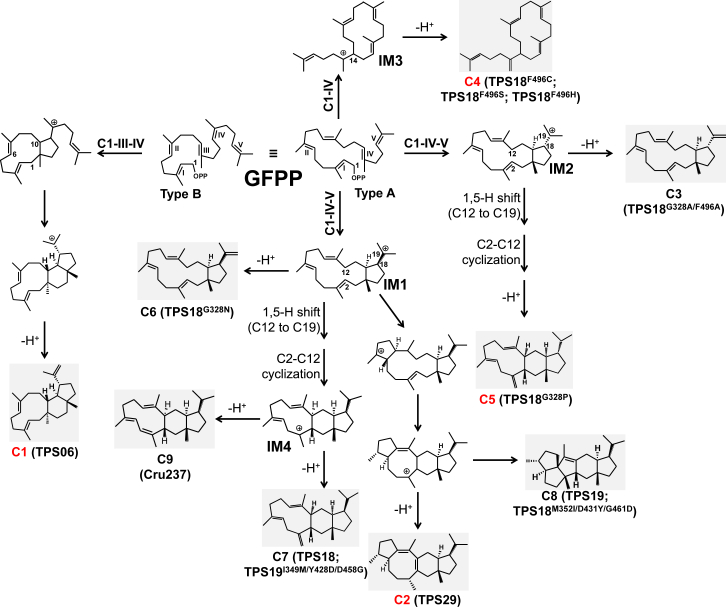
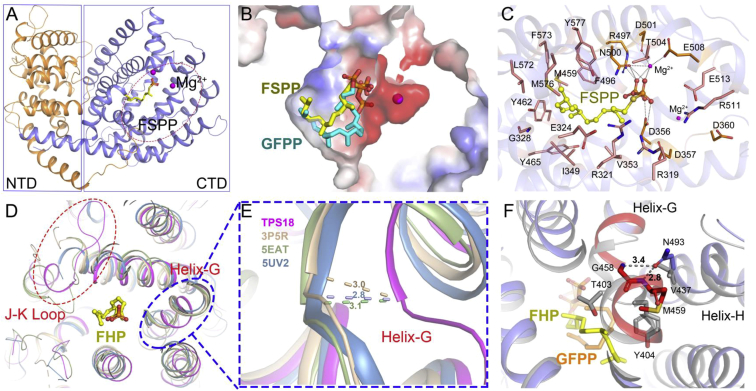
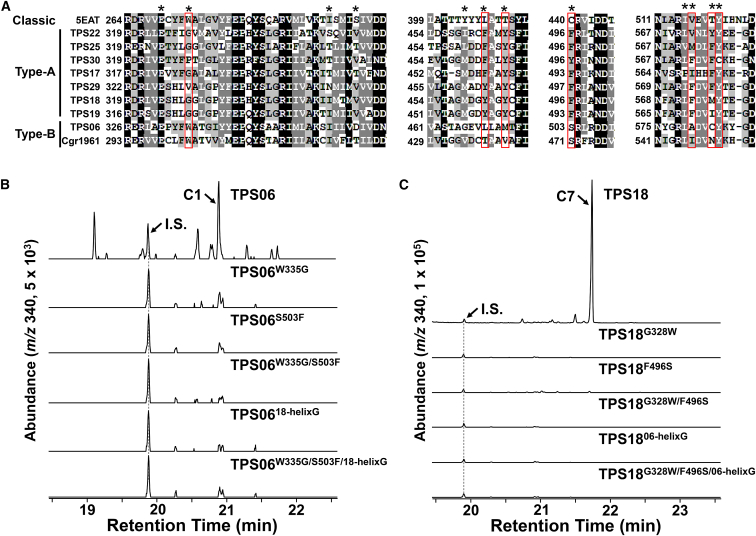
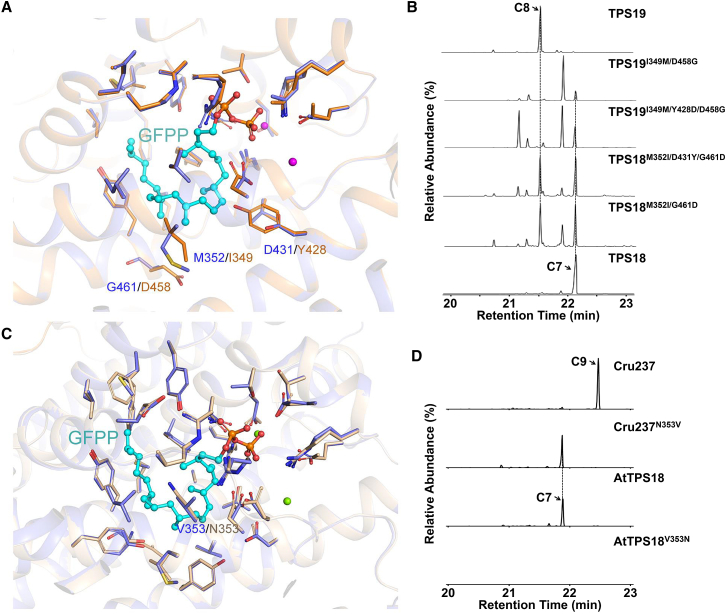
Class I terpene synthase (TPS) generates bioactive terpenoids with diverse backbones. Sesterterpene synthase (sester-TPS, C25), a branch of class I TPSs, was recently identified in Brassicaceae. However, the catalytic mechanisms of sester-TPSs are not fully understood. Here, we first identified three nonclustered functional sester-TPSs (AtTPS06, AtTPS22, and AtTPS29) in Arabidopsis thaliana. AtTPS06 utilizes a type-B cyclization mechanism, whereas most other sester-TPSs produce various sesterterpene backbones via a type-A cyclization mechanism. We then determined the crystal structure of the AtTPS18-FSPP complex to explore the cyclization mechanism of plant sester-TPSs. We used structural comparisons and site-directed mutagenesis to further elucidate the mechanism: (1) mainly due to the outward shift of helix G, plant sester-TPSs have a larger catalytic pocket than do mono-, sesqui-, and di-TPSs to accommodate GFPP; (2) type-A sester-TPSs have more aromatic residues (five or six) in their catalytic pocket than classic TPSs (two or three), which also determines whether the type-A or type-B cyclization mechanism is active; and (3) the other residues responsible for product fidelity are determined by interconversion of AtTPS18 and its close homologs. Altogether, this study improves our understanding of the catalytic mechanism of plant sester-TPS, which ultimately enables the rational engineering of sesterterpenoids for future applications.
Keywords: crystal structure; cyclization mechanism; sesterterpene; terpene synthase; terpenoid.
© 2020 The Author(s).
Figures
Comment in
-
A New Year's spotlight on two years of publication.Plant Commun. 2021 Dec 29;3(1):100274. doi: 10.1016/j.xplc.2021.100274. eCollection 2022 Jan 10. Plant Commun. 2021. PMID: 35059635 Free PMC article. No abstract available.
References
-
- Bian G.K., Han Y.C., Hou A.W., Yuan Y.J., Liu X.H., Deng Z.X., Liu T.G. Releasing the potential power of terpene synthases by a robust precursor supply platform. Metab. Eng. 2017;42:1–8. - PubMed
-
- Chen F., Tholl D., Bohlmann J., Pichersky E. The family of terpene synthases in plants: a mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011;66:212–229. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
