

IFT54 directly interacts with kinesin-II and IFT dynein to regulate anterograde intraflagellar transport
- PMID: 33368450
- PMCID: PMC7917553
- DOI: 10.15252/embj.2020105781
IFT54 directly interacts with kinesin-II and IFT dynein to regulate anterograde intraflagellar transport
Abstract
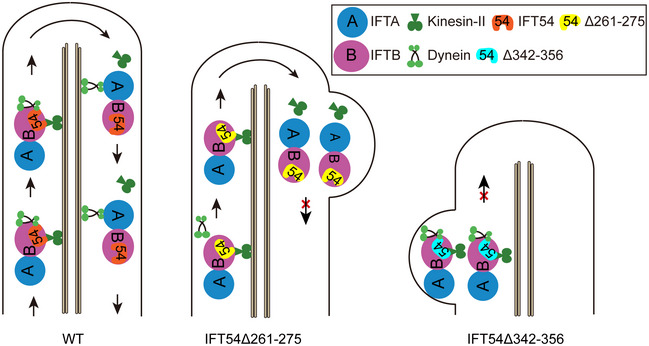
The intraflagellar transport (IFT) machinery consists of the anterograde motor kinesin-II, the retrograde motor IFT dynein, and the IFT-A and -B complexes. However, the interaction among IFT motors and IFT complexes during IFT remains elusive. Here, we show that the IFT-B protein IFT54 interacts with both kinesin-II and IFT dynein and regulates anterograde IFT. Deletion of residues 342-356 of Chlamydomonas IFT54 resulted in diminished anterograde traffic of IFT and accumulation of IFT motors and complexes in the proximal region of cilia. IFT54 directly interacted with kinesin-II and this interaction was strengthened for the IFT54Δ342-356 mutant in vitro and in vivo. The deletion of residues 261-275 of IFT54 reduced ciliary entry and anterograde traffic of IFT dynein with accumulation of IFT complexes near the ciliary tip. IFT54 directly interacted with IFT dynein subunit D1bLIC, and deletion of residues 261-275 reduced this interaction. The interactions between IFT54 and the IFT motors were also observed in mammalian cells. Our data indicate a central role for IFT54 in binding the IFT motors during anterograde IFT.
Keywords: IFT dynein; IFT54; cilia; flagella; intraflagellar transport; kinesin-II.
© 2020 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
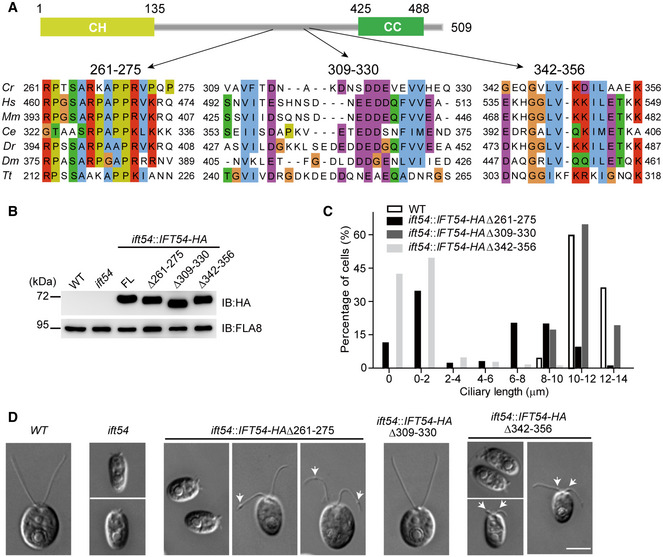
IFT54 has three previously undefined and conserved regions. Schematic representation of the primary structure of Chlamydomonas IFT54 is shown on the top. Three conserved regions are shown on the bottom. Abbreviations for the species and protein entries: Cr, Chlamydomonas reinhardtii (Cre11.g467739); Hs, Homo sapiens (NP_056465.2); Mm, Mus musculus (NP_082994.1); Ce, Caenorhabditis (NP_508149.1); Dr, Danio rerio (ABW96885); Dm, Drosophila melanogaster (NP_650353); Tt, Tetrahymena thermophila (XP_001026831.1).
Immunoblot analysis of whole cell samples of the HA‐tagged full‐length (FL) IFT54, and its deletion mutants as indicated, each expressed in the ift54 null mutant; wild‐type (WT) and ift54 mutant cells were used as control. Anti‐HA was use to visualize the transgenic proteins, and antibodies to FLA8/KIF3B were used as a loading control.
Histogram showing ciliary length distribution in populations of ift54 mutant cells transformed with the deletion mutant constructs as indicated. Wild‐type (WT) cells were used as a control.
Representative DIC images of wild‐type (WT), ift54, and ift54 cells transformed with the deletion constructs as indicated. Arrows indicate cilia bulges. Scale bar, 5 µm.
- A
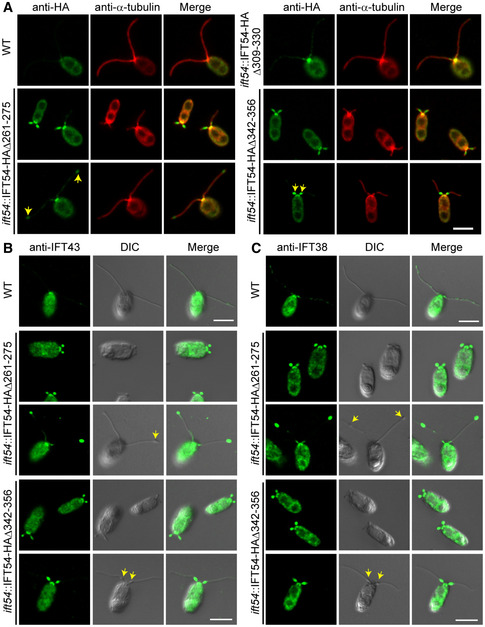
Immunostaining of IFT54 mutated proteins in various deletion mutants as indicated; wild‐type (WT) cells were used as control. Specimens were stained with anti‐HA and anti‐α‐tubulin antibodies followed by imaging using an epifluorescence microscope. Arrows indicate protein accumulation at ciliary tip (left) or proximal end of cilia (right). Scale bars, 5 µm.
- B, C
Immunostaining analysis of IFT43 and IFT38 in the deletion mutants as indicated. Cells were fixed and stained with antibodies against IFT43 (an IFT‐A subunit) (B) or against IFT38 (an IFT‐B subunit) (C) followed by imaging using both DIC and epifluorescence microscopy. IFT43 and IFT38 accumulated at the ciliary tip in IFT54Δ261–275 mutant but at proximal end of cilia in IFT54Δ342–356 mutant. Arrows indicate ciliary bulges. WT, wild‐type cells. Scale bars, 5 µm.
- A
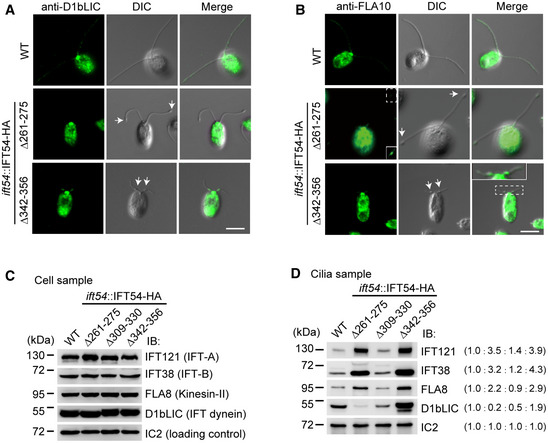
Immunostaining of cells with antibody against IFT dynein subunit D1bLIC. D1bLIC is absent in the cilia bulges at the ciliary tip in IFT54Δ261–275 mutant while enriched at the cilia bulges at proximal end of cilia in IFT54Δ342–356 mutant. WT, wild‐type cells. Arrows indicate ciliary bulges. Scale bar, 5 µm.
- B
Immunostaining of cells with antibody against kinesin‐II subunit FLA10. FLA10 is enriched in cilia bulges at the ciliary tip in IFT54Δ261–275 mutant or at proximal end of cilia in IFT54Δ342–356 mutant. WT, wild‐type cells. The insets show enhanced and/or enlarged images. Arrows indicate ciliary bulges. Scale bar, 5 µm.
- C, D
Immunoblot analysis of whole cells (C) and isolated cilia (D) of wild‐type and the three IFT54 deletion mutants. The membranes were probed with antibodies against IFT complex and motor proteins as indicated. Wild‐type levels of the various IFT components were detected in the whole cell samples of the deletion strains (C). IFT121, IFT38, and FLA8 accumulated in cilia of both IFT54Δ261–275 and IFT54Δ342–356 mutants. D1bLIC was relatively reduced in IFT54Δ261–275 mutant cilia but was substantially increased in IFT54Δ342–356 mutant cilia (D). Ratios of protein amounts from one representative experiment are in parentheses.
- A
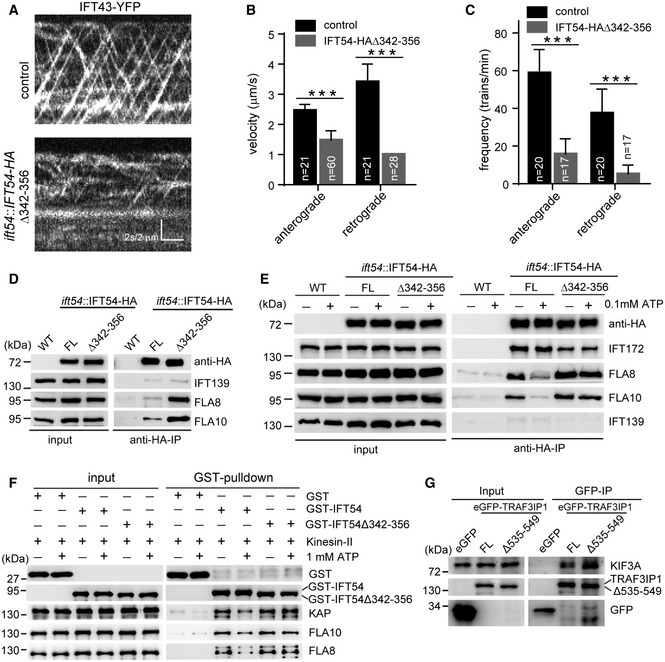
Kymograms showing the trajectories of IFT trains inside cilia as visualized with IFT43‐YFP. An ift43 rescue strain expressing IFT43‐YFP (as a control) and the IFT54Δ342–356 mutant expressing IFT43‐YFP were analyzed by live imaging via TIRF microscopy.
- B, C
Velocities (B) and frequencies (C) of anterograde and retrograde IFT of IFT43‐YFP in control and the IFT54Δ342–356 mutant. n represents the number of cilia assayed from three independent experiments. Values show the mean ± SD. Unpaired two‐tailed Student’s t‐test analysis, ***P < 0.0001.
- D
Deletion of residues 342–356 of IFT54 increases its interaction with kinesin‐II. Cell extracts from wild‐type (WT), ift54 expressing HA‐tagged full‐length (FL) IFT54 or IFT54Δ342–356 mutant were subjected to immunoprecipitation with anti‐HA antibody followed by immunoblotting with the indicated antibodies. The normalized ratios of FLA8 and FLA10 in the immunoprecipitates (IFT54 versus the mutant) are 1:3.90 and 1:3.13, respectively.
- E
ATP treatment induces dissociation of IFT54 but not IFT54Δ342–356 mutant from kinesin‐II. Immunoprecipitation experiments were performed as shown in (D) in the presence or absence of ATP. The normalized ratios of FLA8 and FLA10 in the immunoprecipitates (IFT54 without ATP/with ATP/mutant without ATP/with ATP) are 1:0.37:1.70:1.38 and 1:0.35:1.47:1.45, respectively.
- F
Analysis of the interaction of IFT54 and IFT54Δ342–356 mutant with kinesin‐II by GST pull‐down assay. Bacterial expressed GST, GST‐tagged IFT54, or IFT54Δ342–356 mutant was mixed, respectively, with recombinant kinesin‐II purified from insect cells in the presence or absence of ATP followed by GST pull‐down and immunoblotting. The normalized ratios of KAP, FLA10, and FLA8 in the pull‐down samples (GST‐IFT54 without ATP/with ATP/mutant without ATP/with ATP) are 1:0.51:1.31:1.44, 1:0.55:1.29:1.19, and 1:0.50:1.31:1.29, respectively.
- G
Interaction of IFT54 with kinesin‐II is conserved in mammalian cells. Cell extracts from HEK293T cells expressing eGFP, eGFP‐tagged TRAF3IP1, or TRAF3IP1Δ535–549 (corresponding deletion mutant of IFT54Δ342–356) were analyzed by immunoprecipitation with GFP antibody followed by immunoblotting with anti‐GFP and KIF3A antibodies, respectively. Please note that the deletion mutant had an increased interaction with kinesin‐II subunit KIF3A relative to the control. The normalized ratio of KIF3A in the immunoprecipitates (TRAF3IP1 versus mutant) is 1:1.76.
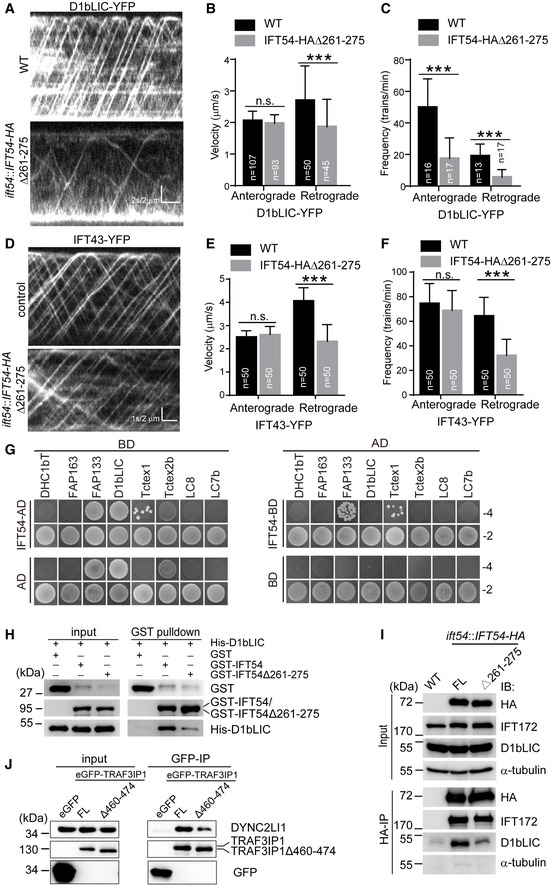
- A
Kymograms showing the trajectories of IFT trains inside cilia as visualized with D1bLIC‐YFP. Wild‐type (WT) and IFT54Δ261–275 mutant cells expressing D1bLIC‐YFP were analyzed by live imaging via TIRF microscopy.
- B, C
Velocities (B) and frequencies (C) of anterograde and retrograde IFT of D1bLIC‐YFP. n represents the number of cilia assayed from four independent experiments. Values show the mean ± SD. Unpaired two‐tailed Student’s t‐test analysis, n.s.: no significance; ***P < 0.0001.
- D
Kymograms of IFT43‐YFP of WT and mutant cilia.
- E, F
Velocities (E) and frequencies (F) of anterograde and retrograde IFT of IFT43‐YFP. n represents the number of cilia assayed from three independent experiments. Values show the mean ± SD. Unpaired two‐tailed Student’s t‐test analysis, n.s.: no significance; ***P < 0.0001.
- G
Yeast two‐hybrid assay for interaction between IFT54 and IFT dynein subunits. Yeast cells that were transformed with each pair of constructs as indicated were grown under selection media lacking leucine, tryptophan, histidine, and adenine (−4) or lacking leucine and tryptophan (−2). DHC1bT, DHC1b tail domain; Empty AD or BD vectors were used as control.
- H
Interaction of IFT54 and IFT54Δ261–275 with D1bLIC by GST pull‐down assay. Bacterial expressed GST, GST‐IFT54, or GST‐IFT54Δ261–275 were mixed, respectively, with His‐D1bLIC followed by GST pull‐down and immunoblotting with anti‐GST and anti‐His antibodies. The normalized ratio of His‐D1bLIC in the pull‐down samples (GST‐IFT54 versus mutant) is 1:0.32.
- I
Co‐immunoprecipitation of IFT54 and IFT54Δ261–275 with D1bLIC. Cell extracts from wild‐type (WT), ift54 expressing HA‐tagged full‐length (FL) IFT54 or IFT54Δ261–275 mutant were subjected to immunoprecipitation with anti‐HA antibody followed by immunoblotting with the indicated antibodies. Please note that IFT54Δ261–275 mutant exhibited weaker interaction with D1bLIC relative to the control. The normalized ratio of D1bLIC in the immunoprecipitates (IFT54‐HA versus mutant) is 1:0.37.
- J
Interaction of IFT54 with D1bLIC is conserved in mammalian cells. Cell extracts from HEK293T cells expressing eGFP, eGFP‐tagged TRAF3IP1, or TRAF3IP1Δ460–474 (corresponding deletion mutant of IFT54Δ261–275) were analyzed by immunoprecipitation with GFP antibody followed by immunoblotting with anti‐GFP and DYNC2LI1, human homologue of D1bLIC, antibodies, respectively. Please note that the deletion mutant weakened the interaction with IFT dynein subunit DYNC2LI1 relative to the control. The normalized ratio of DYNC2LI1 in the immunoprecipitates (TRAF3IP1 versus mutant) is 1:0.49.
References
-
- Baker SA, Freeman K, Luby‐Phelps K, Pazour GJ, Besharse JC (2003) IFT20 links kinesin II with a mammalian intraflagellar transport complex that is conserved in motile flagella and sensory cilia. J Biol Chem 278: 34211–34218 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
