

Molecular signatures of sexual communication in the phlebotomine sand flies
- PMID: 33370303
- PMCID: PMC7793272
- DOI: 10.1371/journal.pntd.0008967
Molecular signatures of sexual communication in the phlebotomine sand flies
Abstract
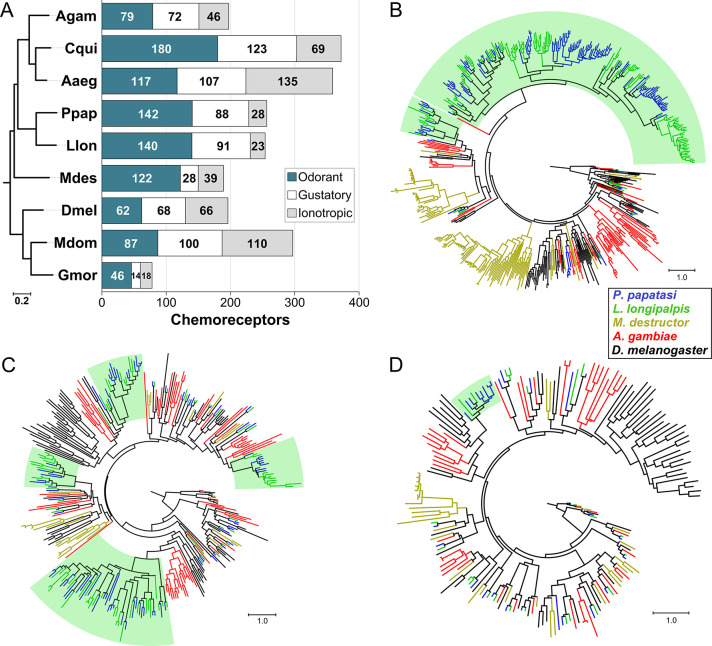
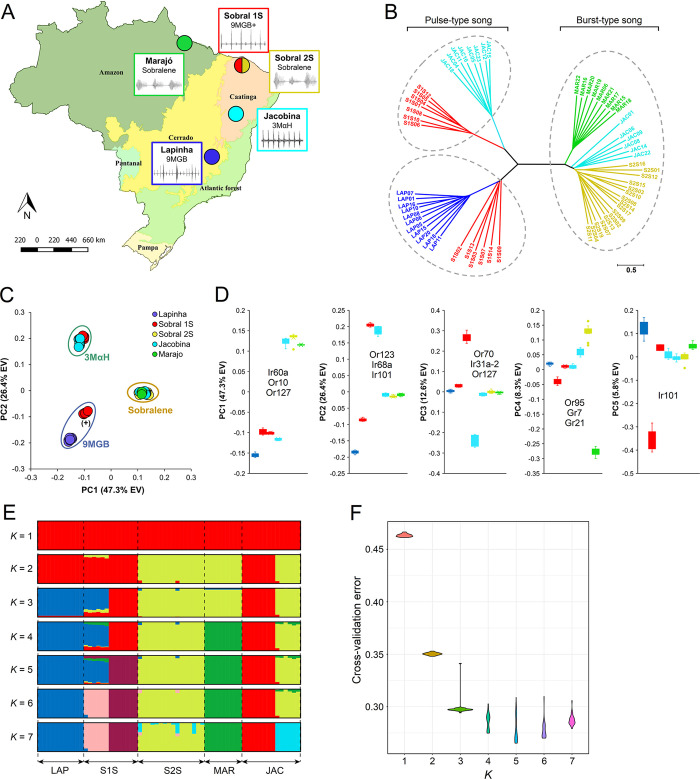
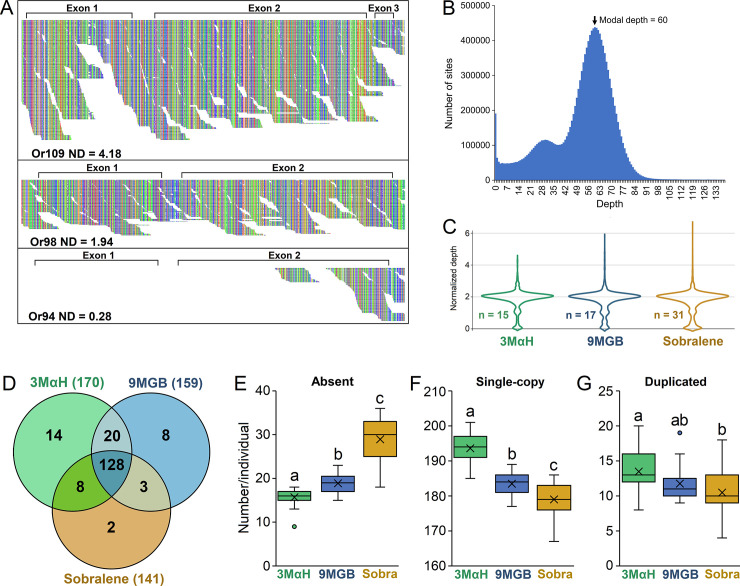
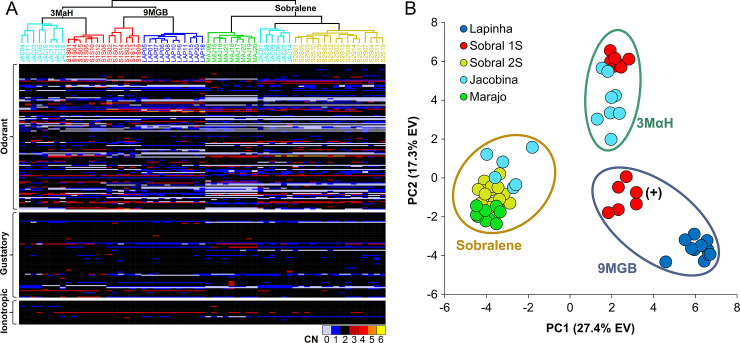
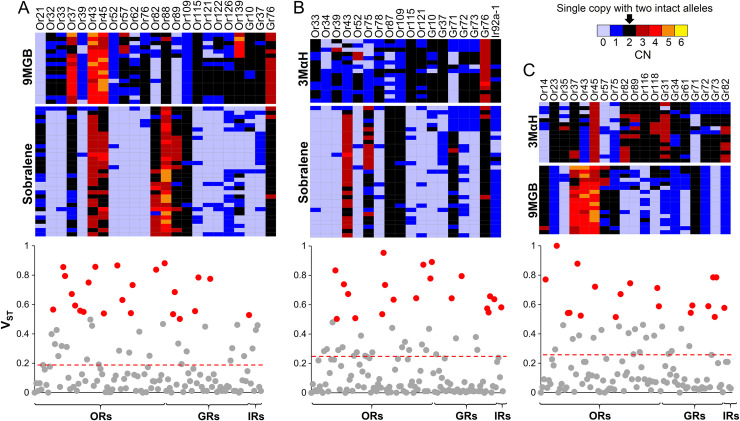
Phlebotomine sand flies employ an elaborate system of pheromone communication wherein males produce pheromones that attract other males to leks (thus acting as an aggregation pheromone) and females to the lekking males (sex pheromone). In addition, the type of pheromone produced varies among populations. Despite the numerous studies on sand fly chemical communication, little is known of their chemosensory genome. Chemoreceptors interact with chemicals in an organism's environment to elicit essential behaviors such as the identification of suitable mates and food sources. Thus, they play important roles during adaptation and speciation. Major chemoreceptor gene families, odorant receptors (ORs), gustatory receptors (GRs) and ionotropic receptors (IRs) together detect and discriminate the chemical landscape. Here, we annotated the chemoreceptor repertoire in the genomes of Lutzomyia longipalpis and Phlebotomus papatasi, major phlebotomine vectors in the New World and Old World, respectively. Comparison with other sequenced Diptera revealed a large and unique expansion where over 80% of the ~140 ORs belong to a single, taxonomically restricted clade. We next conducted a comprehensive analysis of the chemoreceptors in 63 L. longipalpis individuals from four different locations in Brazil representing allopatric and sympatric populations and three sex-aggregation pheromone types (chemotypes). Population structure based on single nucleotide polymorphisms (SNPs) and gene copy number in the chemoreceptors corresponded with their putative chemotypes, and corroborate previous studies that identified multiple populations. Our work provides genomic insights into the underlying behavioral evolution of sexual communication in the L. longipalpis species complex in Brazil, and highlights the importance of accounting for the ongoing speciation in central and South American Lutzomyia that could have important implications for vectorial capacity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- WHO. Leishmaniasis: https://www.who.int/news-room/fact-sheets/detail/leishmanreviewiasis; 2020 [cited 2020].
-
- WHO. Leishmaniasis, The Vector: https://www.who.int/leishmaniasis/disease/vector/en/; 2020.
-
- Bray DP, Ward RD, Hamilton JG. The chemical ecology of sandflies. In: Takken W, Knols B, editors. Ecology and control of vector-borne diseases 2010. p. 203–16.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
