

Toxoplasma gondii PPM3C, a secreted protein phosphatase, affects parasitophorous vacuole effector export
- PMID: 33370417
- PMCID: PMC7793252
- DOI: 10.1371/journal.ppat.1008771
Toxoplasma gondii PPM3C, a secreted protein phosphatase, affects parasitophorous vacuole effector export
Abstract
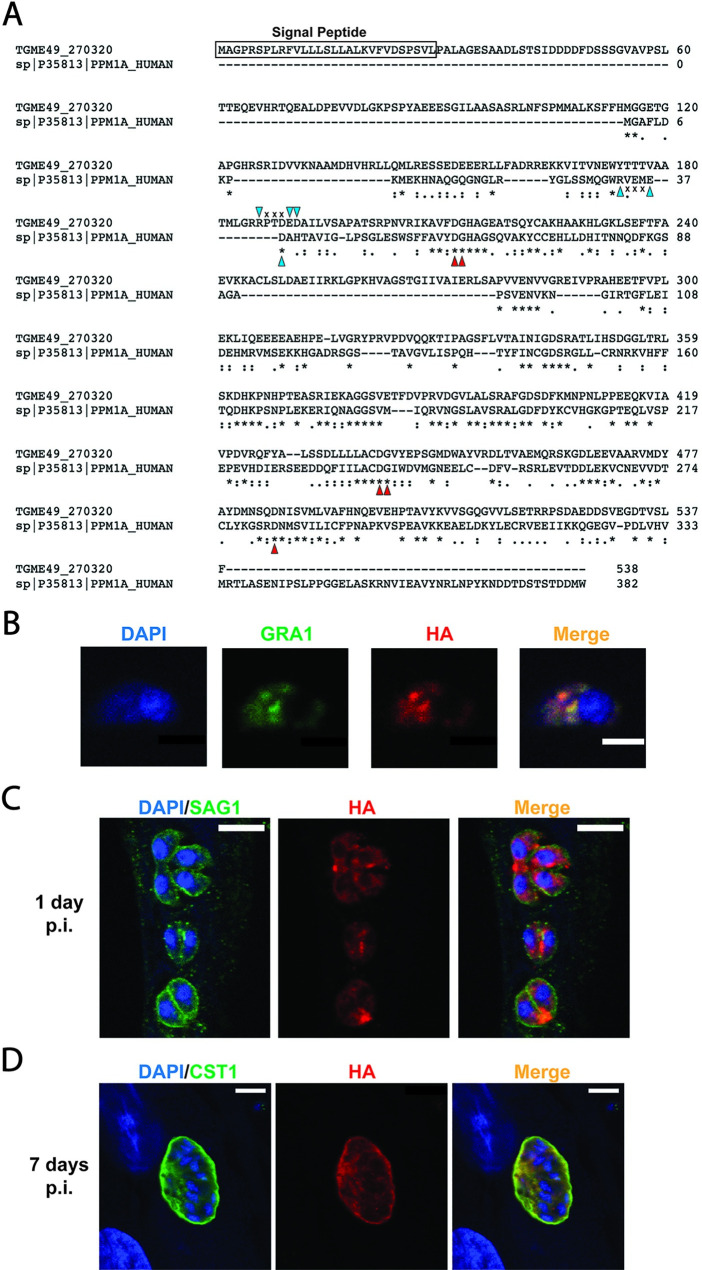
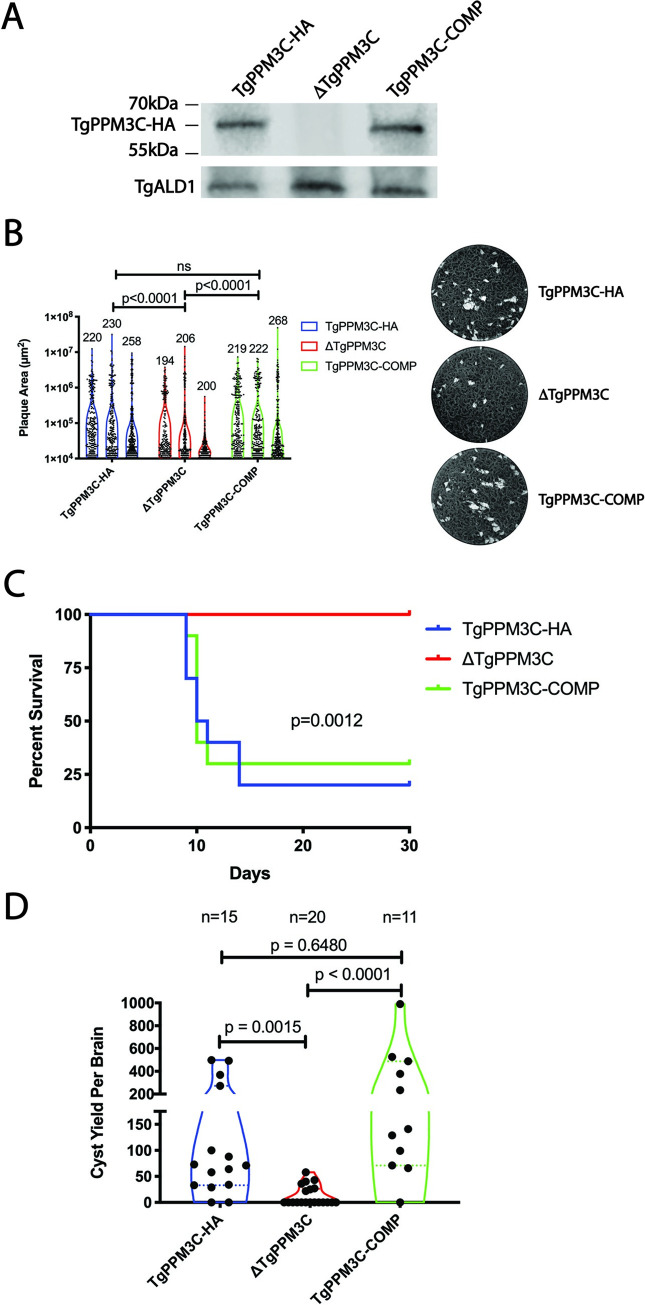
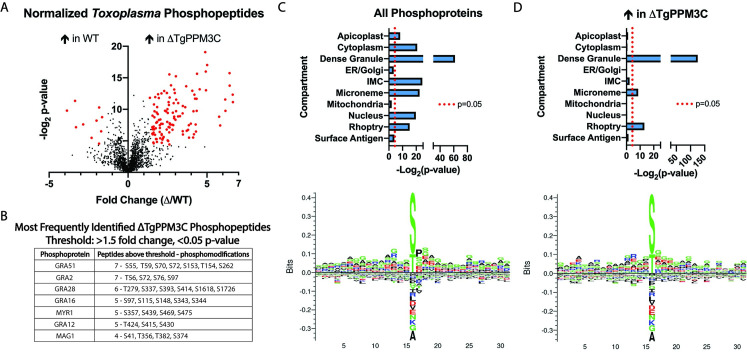
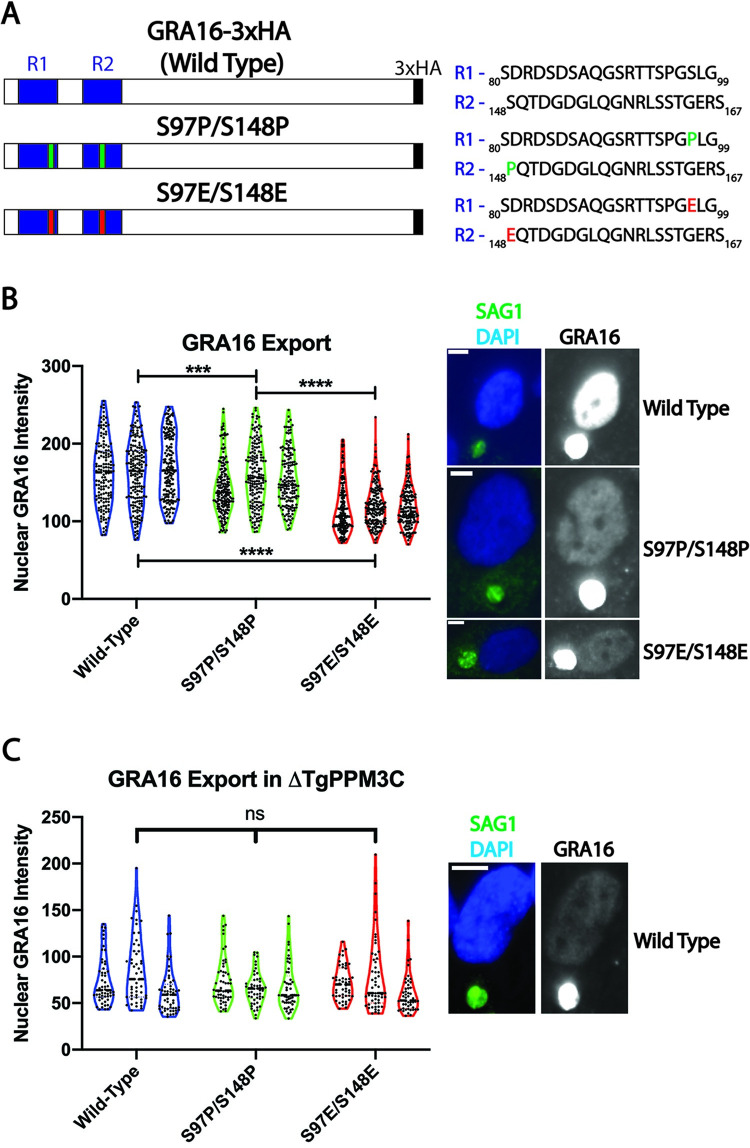
The intracellular parasite Toxoplasma gondii infects a large proportion of humans worldwide and can cause adverse complications in the settings of immune-compromise and pregnancy. T. gondii thrives within many different cell types due in part to its residence within a specialized and heavily modified compartment in which the parasite divides, termed the parasitophorous vacuole. Within this vacuole, numerous proteins optimize intracellular survival following their secretion by the parasite. We investigated the contribution of one of these proteins, TgPPM3C, predicted to contain a PP2C-class serine/threonine phosphatase domain and previously shown to interact with the protein MYR1, an essential component of a putative vacuolar translocon that mediates effector export into the host cell. Parasites lacking the TgPPM3C gene exhibit a minor growth defect in vitro, are avirulent during acute infection in mice, and form fewer cysts in mouse brain during chronic infection. Phosphoproteomic assessment of TgPPM3C deleted parasite cultures demonstrated alterations in the phosphorylation status of many secreted vacuolar proteins including two exported effector proteins, GRA16 and GRA28, as well as MYR1. Parasites lacking TgPPM3C are defective in GRA16 and GRA28 export, but not in the export of other MYR1-dependant effectors. Phosphomimetic mutation of two GRA16 serine residues results in export defects, suggesting that de-phosphorylation is a critical step in the process of GRA16 export. These findings provide another example of the emerging role of phosphatases in regulating the complex environment of the T. gondii parasitophorous vacuole and influencing the export of specific effector proteins from the vacuolar lumen into the host cell.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Coimmunoprecipitation with MYR1 Identifies Three Additional Proteins within the Toxoplasma gondii Parasitophorous Vacuole Required for Translocation of Dense Granule Effectors into Host Cells.mSphere. 2020 Feb 19;5(1):e00858-19. doi: 10.1128/mSphere.00858-19. mSphere. 2020. PMID: 32075880 Free PMC article.
-
Translocation of Dense Granule Effectors across the Parasitophorous Vacuole Membrane in Toxoplasma-Infected Cells Requires the Activity of ROP17, a Rhoptry Protein Kinase.mSphere. 2019 Jul 31;4(4):e00276-19. doi: 10.1128/mSphere.00276-19. mSphere. 2019. PMID: 31366709 Free PMC article.
-
A Novel Secreted Protein, MYR1, Is Central to Toxoplasma's Manipulation of Host Cells.mBio. 2016 Feb 2;7(1):e02231-15. doi: 10.1128/mBio.02231-15. mBio. 2016. PMID: 26838724 Free PMC article.
-
Crossing the Vacuolar Rubicon: Structural Insights into Effector Protein Trafficking in Apicomplexan Parasites.Microorganisms. 2020 Jun 8;8(6):865. doi: 10.3390/microorganisms8060865. Microorganisms. 2020. PMID: 32521667 Free PMC article. Review.
-
Toxoplasma Mechanisms for Delivery of Proteins and Uptake of Nutrients Across the Host-Pathogen Interface.Annu Rev Microbiol. 2020 Sep 8;74:567-586. doi: 10.1146/annurev-micro-011720-122318. Epub 2020 Jul 17. Annu Rev Microbiol. 2020. PMID: 32680452 Free PMC article. Review.
Cited by
-
Toxoplasma protein export and effector function.Nat Microbiol. 2024 Jan;9(1):17-28. doi: 10.1038/s41564-023-01563-z. Epub 2024 Jan 3. Nat Microbiol. 2024. PMID: 38172621 Review.
-
Transport mechanisms at the malaria parasite-host cell interface.PLoS Pathog. 2021 Apr 1;17(4):e1009394. doi: 10.1371/journal.ppat.1009394. eCollection 2021 Apr. PLoS Pathog. 2021. PMID: 33793667 Free PMC article. Review.
-
Secreted Effectors Modulating Immune Responses to Toxoplasma gondii.Life (Basel). 2021 Sep 20;11(9):988. doi: 10.3390/life11090988. Life (Basel). 2021. PMID: 34575137 Free PMC article. Review.
-
Functional Characterization of 17 Protein Serine/Threonine Phosphatases in Toxoplasma gondii Using CRISPR-Cas9 System.Front Cell Dev Biol. 2022 Jan 10;9:738794. doi: 10.3389/fcell.2021.738794. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35083211 Free PMC article.
-
Phosphatase UBLCP1 is required for the growth, virulence and mitochondrial integrity of Toxoplasma gondii.Parasit Vectors. 2025 Mar 28;18(1):122. doi: 10.1186/s13071-025-06766-3. Parasit Vectors. 2025. PMID: 40156024 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
