

Metastasis and Immune Evasion from Extracellular cGAMP Hydrolysis
- PMID: 33372007
- PMCID: PMC8102348
- DOI: 10.1158/2159-8290.CD-20-0387
Metastasis and Immune Evasion from Extracellular cGAMP Hydrolysis
Abstract
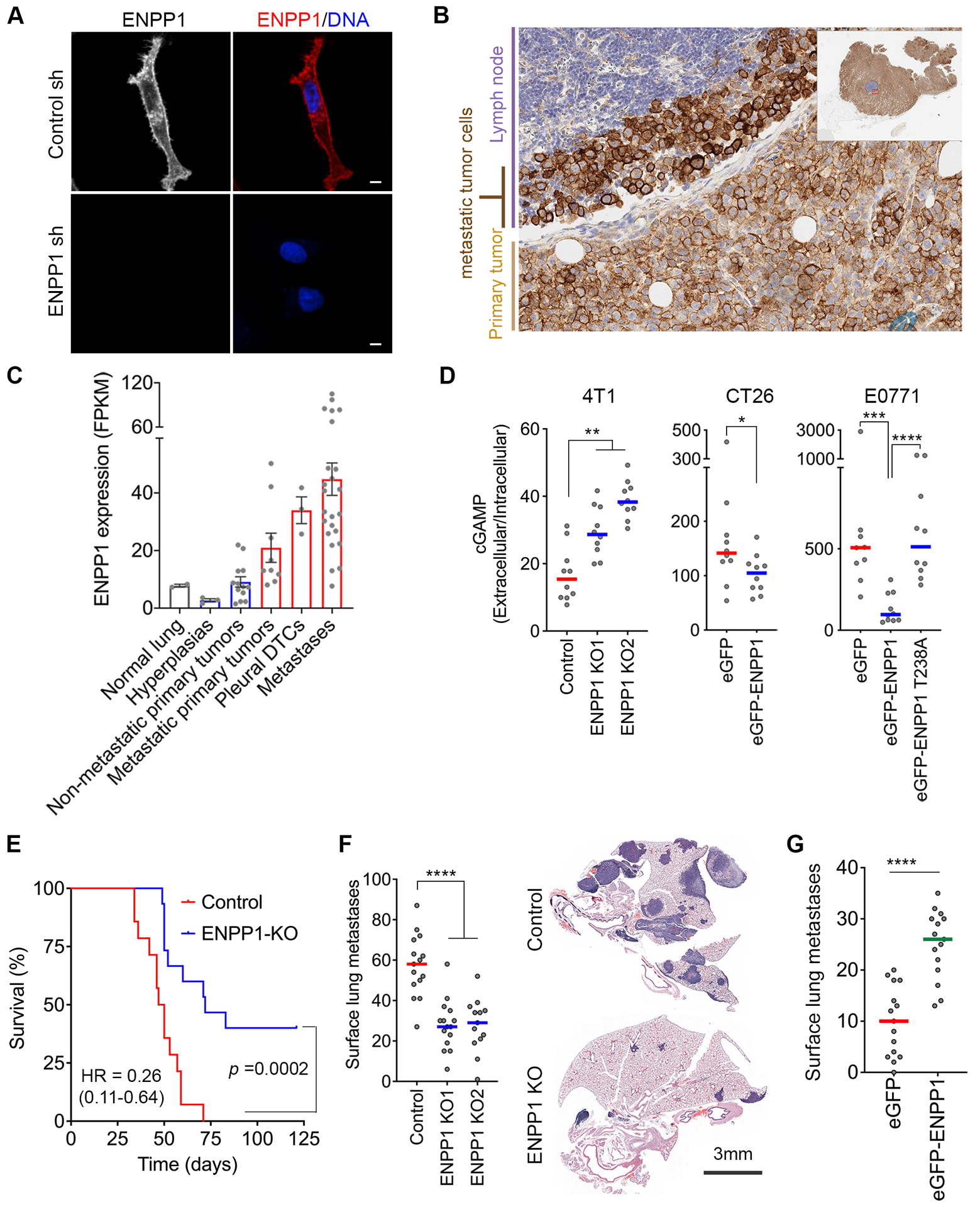
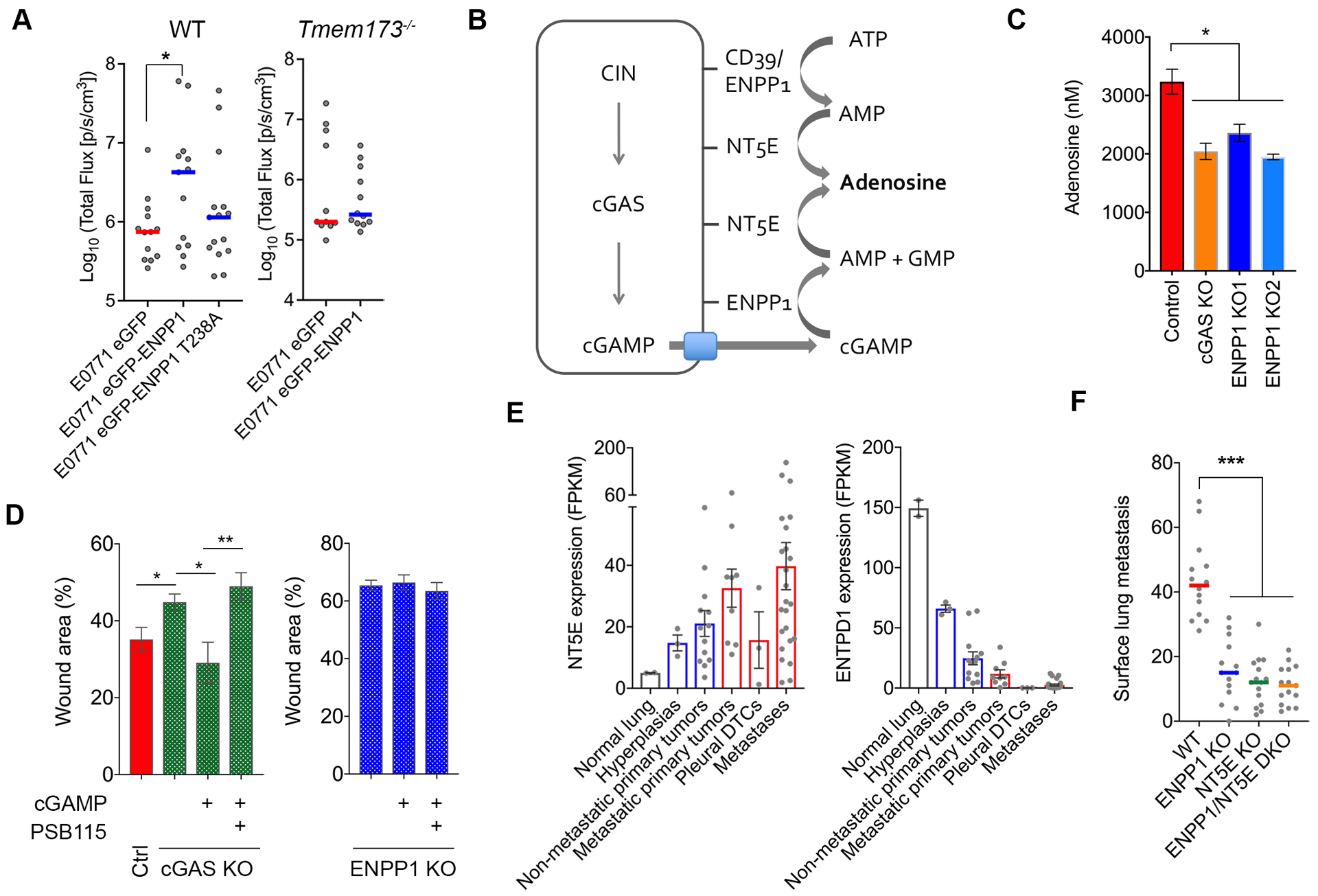
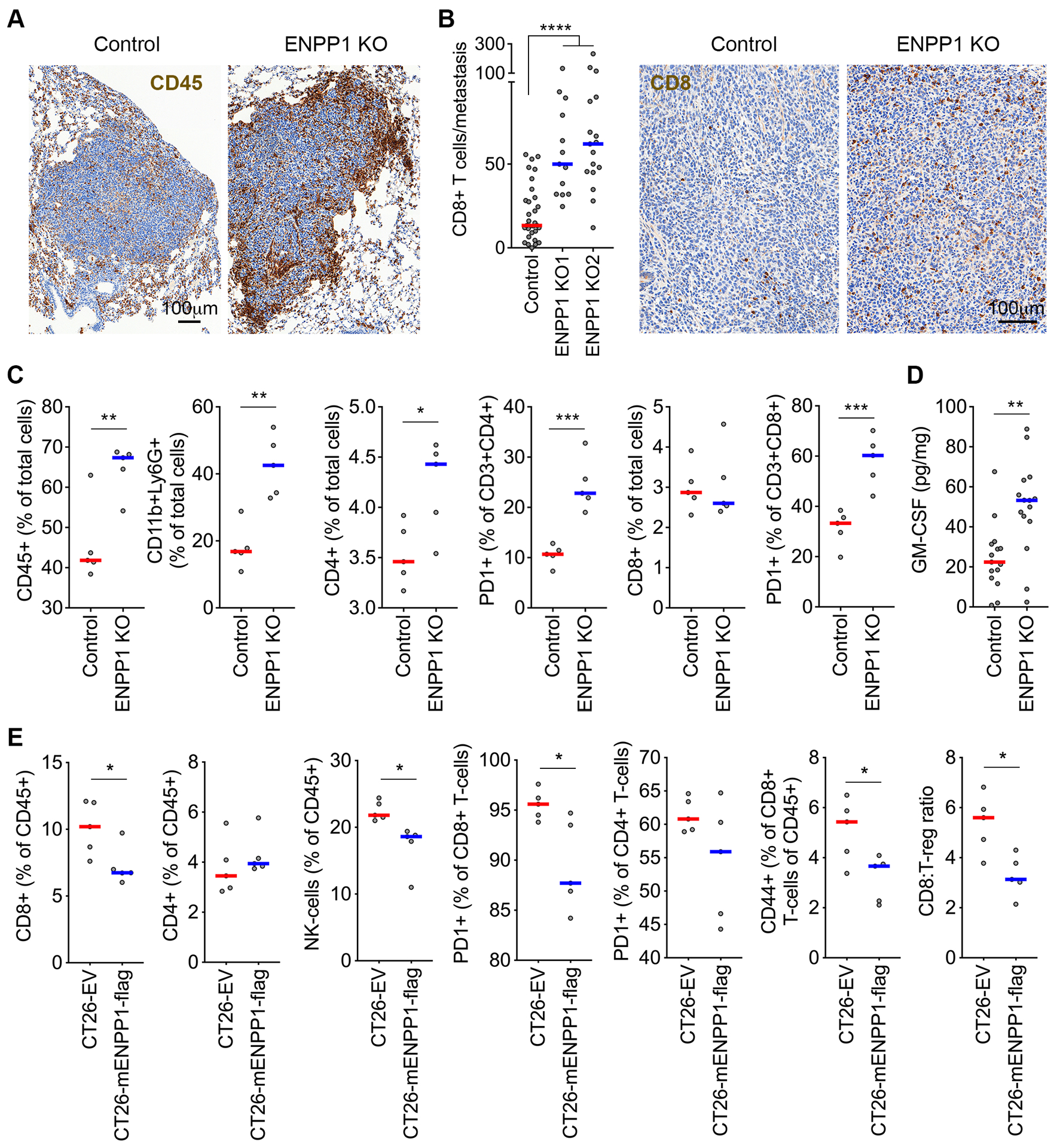
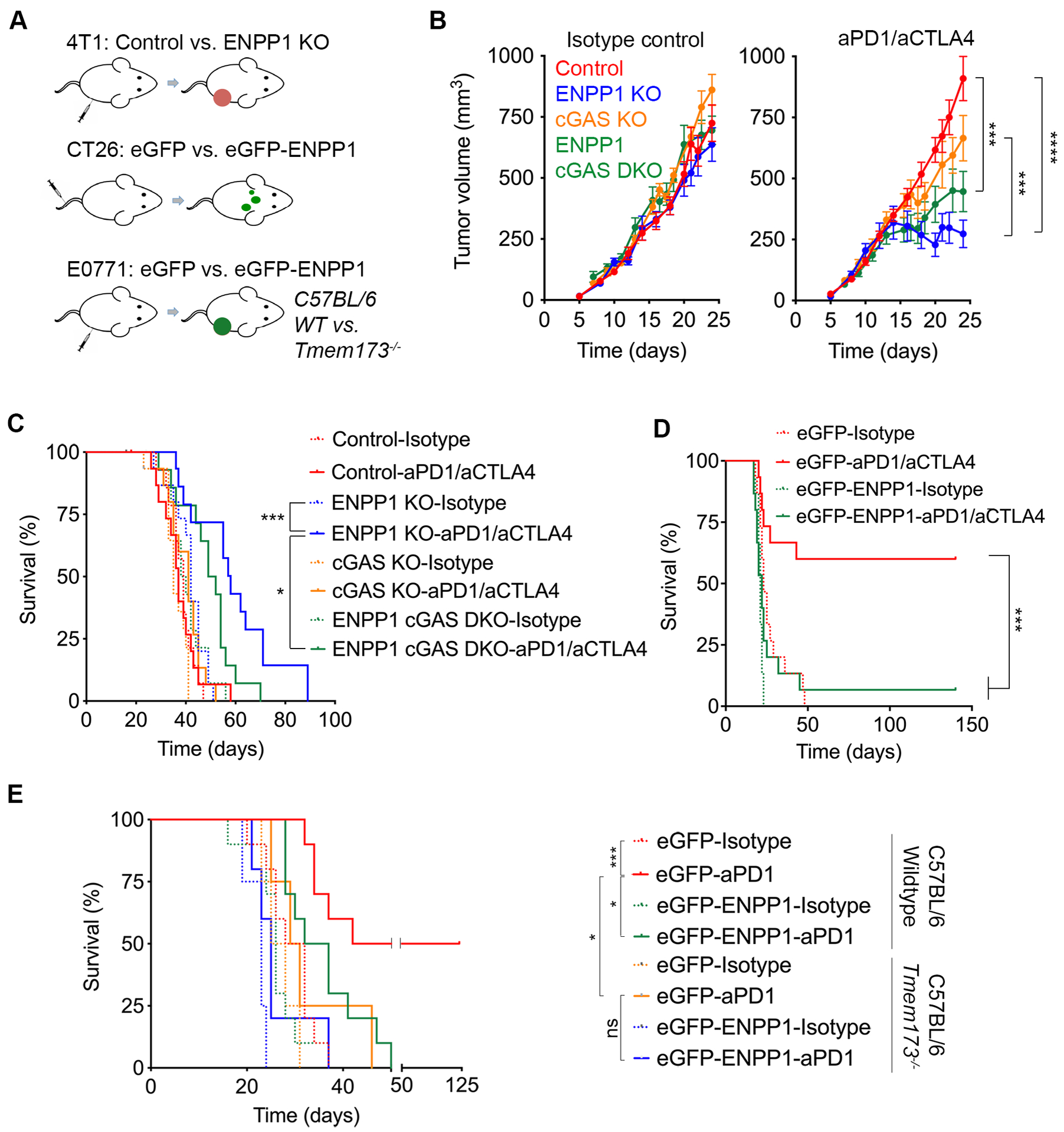
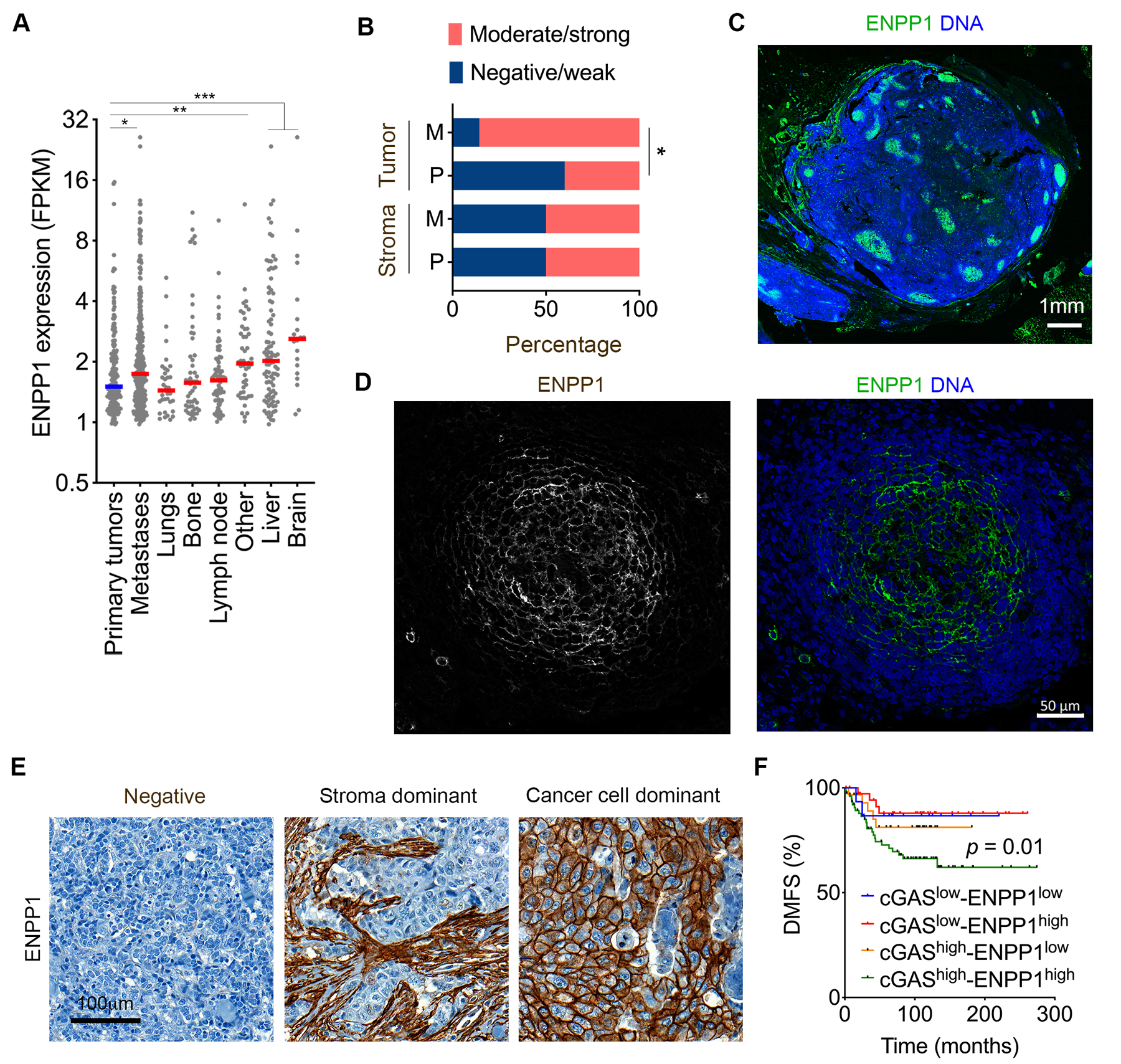
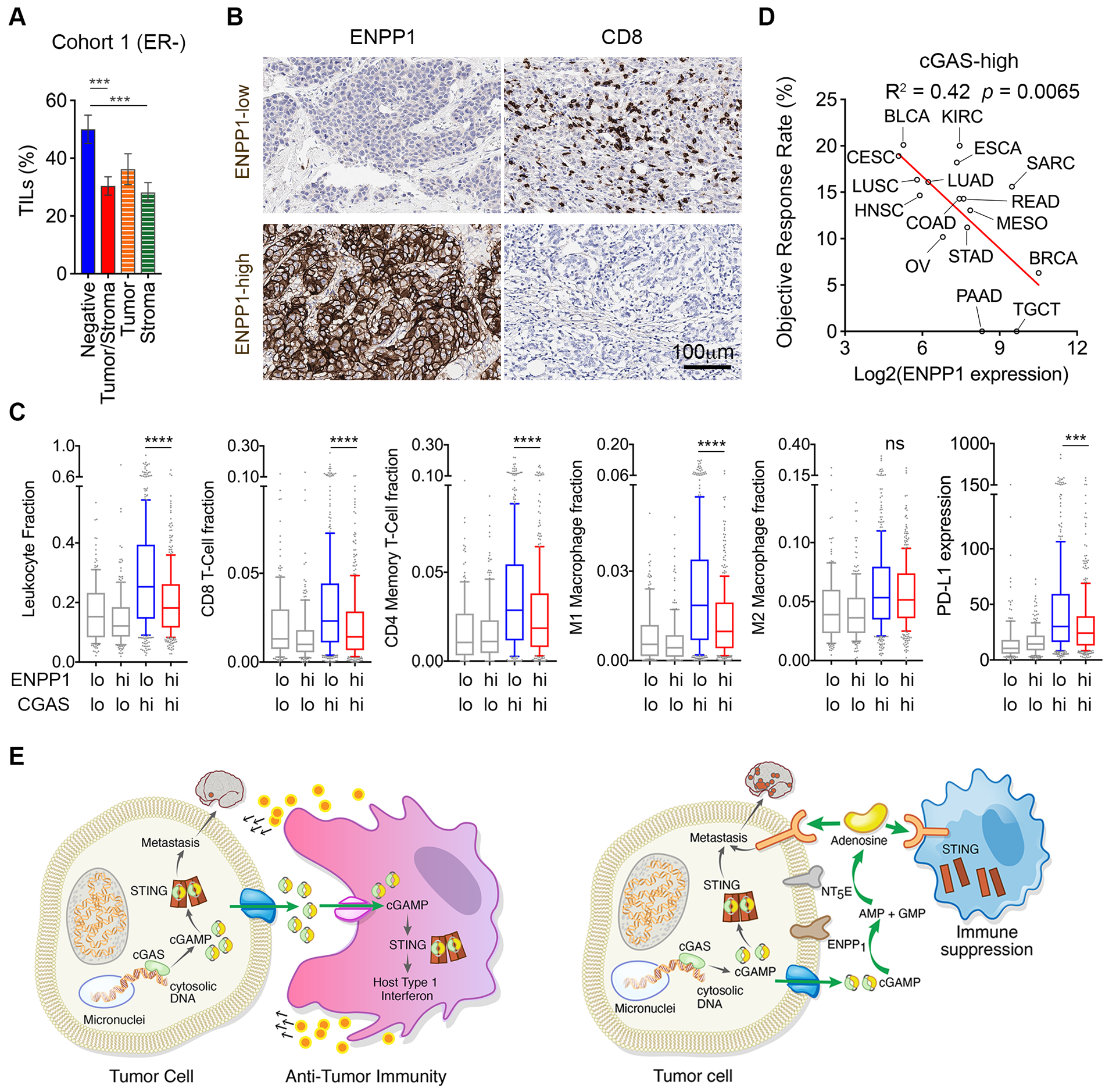
Cytosolic DNA is characteristic of chromosomally unstable metastatic cancer cells, resulting in constitutive activation of the cGAS-STING innate immune pathway. How tumors co-opt inflammatory signaling while evading immune surveillance remains unknown. Here, we show that the ectonucleotidase ENPP1 promotes metastasis by selectively degrading extracellular cGAMP, an immune-stimulatory metabolite whose breakdown products include the immune suppressor adenosine. ENPP1 loss suppresses metastasis, restores tumor immune infiltration, and potentiates response to immune checkpoint blockade in a manner dependent on tumor cGAS and host STING. Conversely, overexpression of wild-type ENPP1, but not an enzymatically weakened mutant, promotes migration and metastasis, in part through the generation of extracellular adenosine, and renders otherwise sensitive tumors completely resistant to immunotherapy. In human cancers, ENPP1 expression correlates with reduced immune cell infiltration, increased metastasis, and resistance to anti-PD-1/PD-L1 treatment. Thus, cGAMP hydrolysis by ENPP1 enables chromosomally unstable tumors to transmute cGAS activation into an immune-suppressive pathway. SIGNIFICANCE: Chromosomal instability promotes metastasis by generating chronic tumor inflammation. ENPP1 facilitates metastasis and enables tumor cells to tolerate inflammation by hydrolyzing the immunotransmitter cGAMP, preventing its transfer from cancer cells to immune cells.This article is highlighted in the In This Issue feature, p. 995.
©2020 American Association for Cancer Research.
Conflict of interest statement
CONFLICTS OF INTEREST STATEMENT
SFB holds a patent related to some of the work described targeting CIN and the cGAS-STING pathway in advanced cancer. He owns equity in, receives compensation from, and serves as a consultant and the Scientific Advisory Board and Board of Directors of Volastra Therapeutics Inc. He has also consulted for Sanofi, received sponsored travel from the Prostate Cancer Foundation, and both travel and compensation from Cancer Research UK. JDW served as a consultant for Adaptive Biotech, Advaxis, Amgen, Apricity, Array BioPharma, Ascentage Pharma, Astellas, Bayer, Beigene, Bristol Myers Squibb, Celgene, Chugai, Elucida, Eli Lilly, F Star, Genentech, Imvaq, Janssen, Kleo Pharma, Kyowa Hakko Kirin, Linneaus, MedImmune, Merck, Neon Therapuetics, Northern Biologics, Ono, Polaris Pharma, Polynoma, Psioxus, Puretech, Recepta, Takara Bio, Trieza, Sellas Life Sciences, Serametrix, Surface Oncology, Syndax, and Syntalogic. He also receives research support from Bristol Myers Squibb, Medimmune, Merck Pharmaceuticals, and Genentech. He owns equity in Potenza Therapeutics, Tizona Pharmaceuticals, Adaptive Biotechnologies, Elucida, Imvaq, Beigene, Trieza, and Linneaus. He has received honoraium from Esanex. TM is a consultant for Immunos Therapeutics and Pfizer; is a co-founder with equity in IMVAQ therapeutics; receives research funding from Bristol-Myers Squibb, Surface Oncology, Kyn Therapeutics, Infinity Pharmaceuticals Inc., Peregrine Pharmaceuticals Inc., Adaptive Biotechnologies, Leap Therapeutics Inc., and Aprea; is an inventor on patent applications related to work on Oncolytic Viral therapy, Alpha Virus Based Vaccine, Neo Antigen Modeling, CD40, GITR, OX40, PD-1 and CTLA-4. KL reports speaker fees from Roche Tissue Diagnostics, travel compensation from BMS and grant income for Genetech. JSR-F has received fees for consulting for Goldman Sachs, REPARE Therapeutics and Paige.AI, and serves as an advisory board member for Roche Diagnostics, InVicro, Genentech, Paige.AI, Volition RX, REPARE Therapeutics and GRAIL. C.S. receives grant support from Pfizer, AstraZeneca, BMS, Roche-Ventana and Boehringer-Ingelheim. C.S. has consulted for Pfizer, Novartis, GlaxoSmithKline, MSD, BMS, Celgene, AstraZeneca, Illumina, Genentech, Roche-Ventana, GRAIL, Medicxi, the Sarah Cannon Research Institute and is an Advisor for Dynamo Therapeutics. C.S. is a shareholder of Apogen Biotechnologies, Epic Bioscience, GRAIL, and has stock options in and is co-founder of Achilles Therapeutics. Outside of the submitted work, K.L. and C.S. have a patent on indel burden and checkpoint inhibitor response pending and a patent on targeting of frameshift neoantigens for personalized immunotherapy pending. BI is a consultant for Merck and Volastra Therapeutics Inc. Remaining authors declare no conflicts of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
