

Pathogenesis of Relapsing Fever
- PMID: 33372163
- PMCID: PMC8756760
- DOI: 10.21775/cimb.042.519
Pathogenesis of Relapsing Fever
Abstract
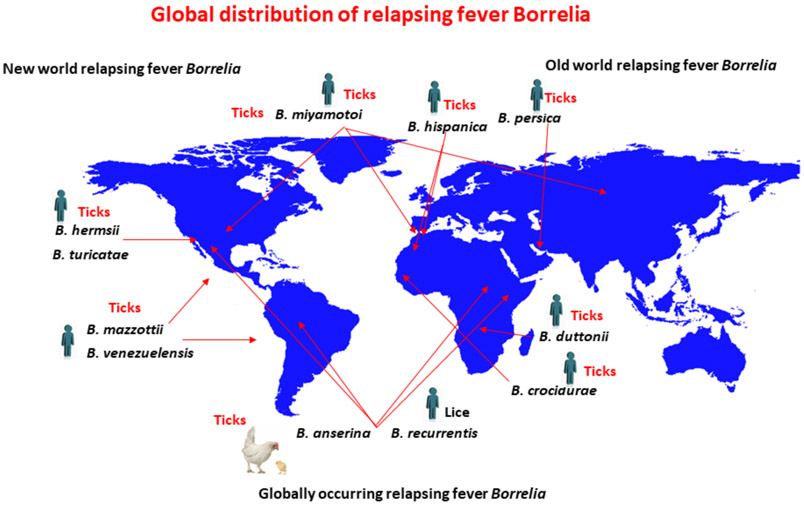
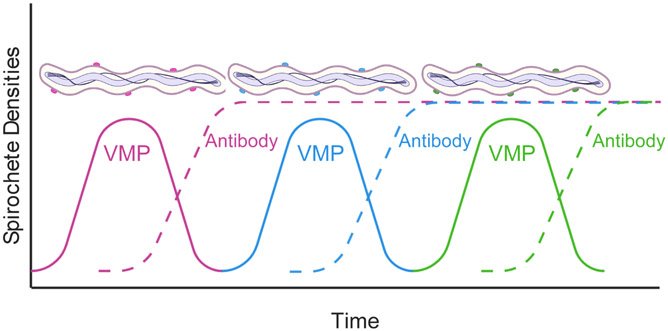
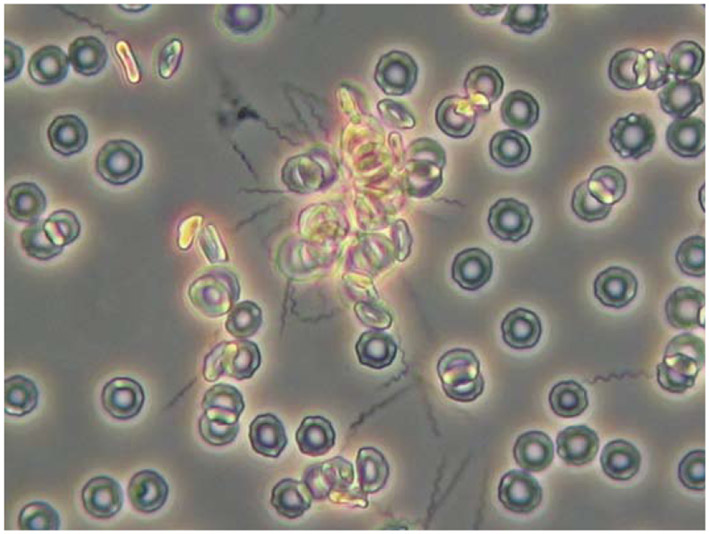
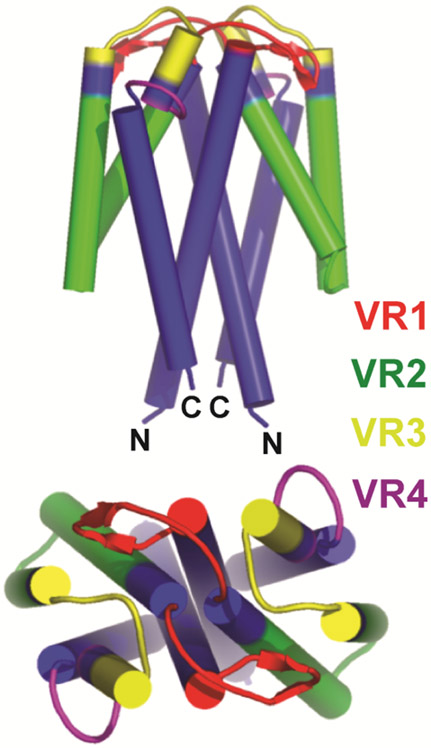
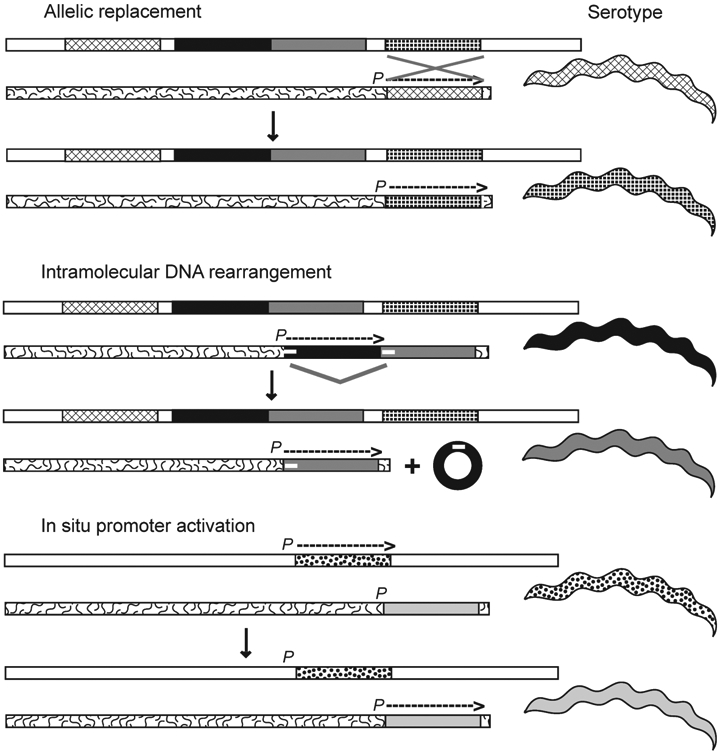
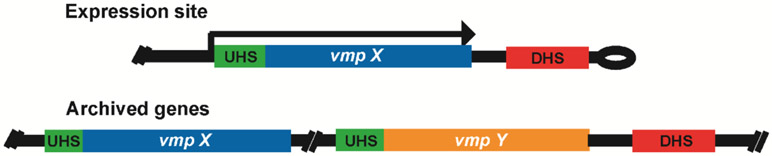
Relapsing fever (RF) is caused by several species of Borrelia; all, except two species, are transmitted to humans by soft (argasid) ticks. The species B. recurrentis is transmitted from one human to another by the body louse, while B. miyamotoi is vectored by hard-bodied ixodid tick species. RF Borrelia have several pathogenic features that facilitate invasion and dissemination in the infected host. In this article we discuss the dynamics of vector acquisition and subsequent transmission of RF Borrelia to their vertebrate hosts. We also review taxonomic challenges for RF Borrelia as new species have been isolated throughout the globe. Moreover, aspects of pathogenesis including symptomology, neurotropism, erythrocyte and platelet adhesion are discussed. We expound on RF Borrelia evasion strategies for innate and adaptive immunity, focusing on the most fundamental pathogenetic attributes, multiphasic antigenic variation. Lastly, we review new and emerging species of RF Borrelia and discuss future directions for this global disease.
Figures
References
-
- Aeschlimann A (1958). Développement embryonnaire d'Ornithodorus moubata (Murray) et transmission transovarienne de Borrelia duttoni. Acta Tropica 15, 15–64. - PubMed
-
- Alugupalli KR, Michelson AD, Barnard MR, Robbins D, Coburn J, Baker EK, Ginsberg MH, Schwan TG, and Leong JM (2001). Platelet activation by a relapsing fever spirochaete results in enhanced bacterium-platelet interaction via integrin alphaIIbbeta3 activation. Mol Microbiol 39, 330–340. doi:10.1046/j.1365-2958.2001.02201.x - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
