

Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) membrane (M) protein inhibits type I and III interferon production by targeting RIG-I/MDA-5 signaling
- PMID: 33372174
- PMCID: PMC7768267
- DOI: 10.1038/s41392-020-00438-7
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) membrane (M) protein inhibits type I and III interferon production by targeting RIG-I/MDA-5 signaling
Abstract
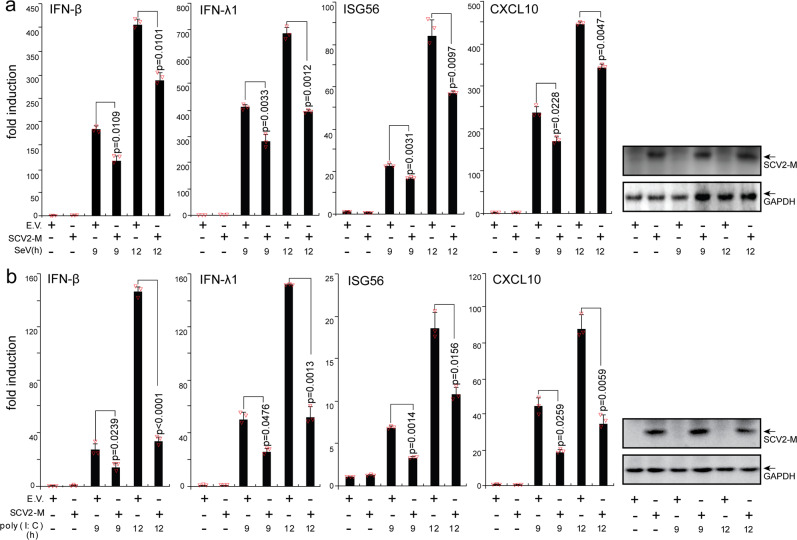
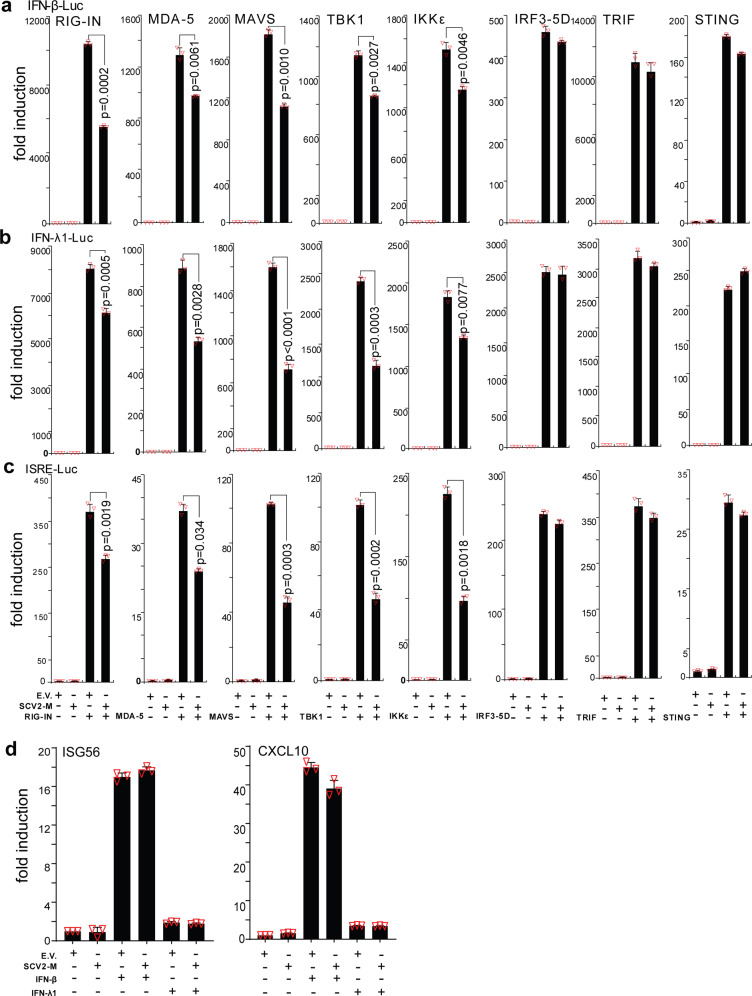
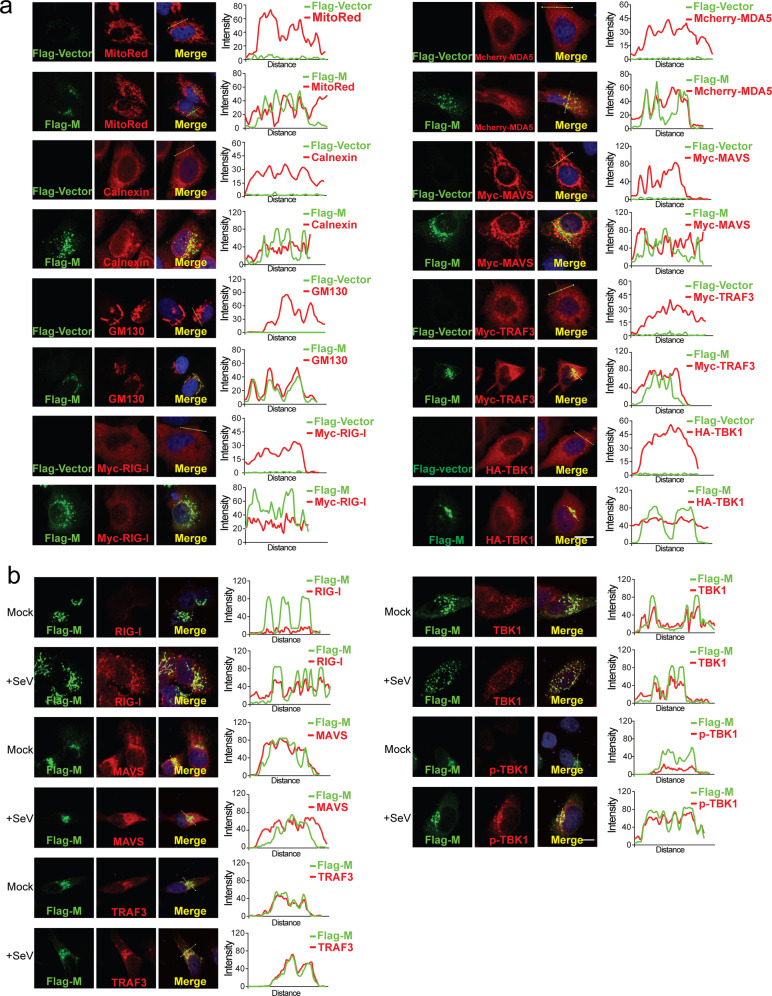
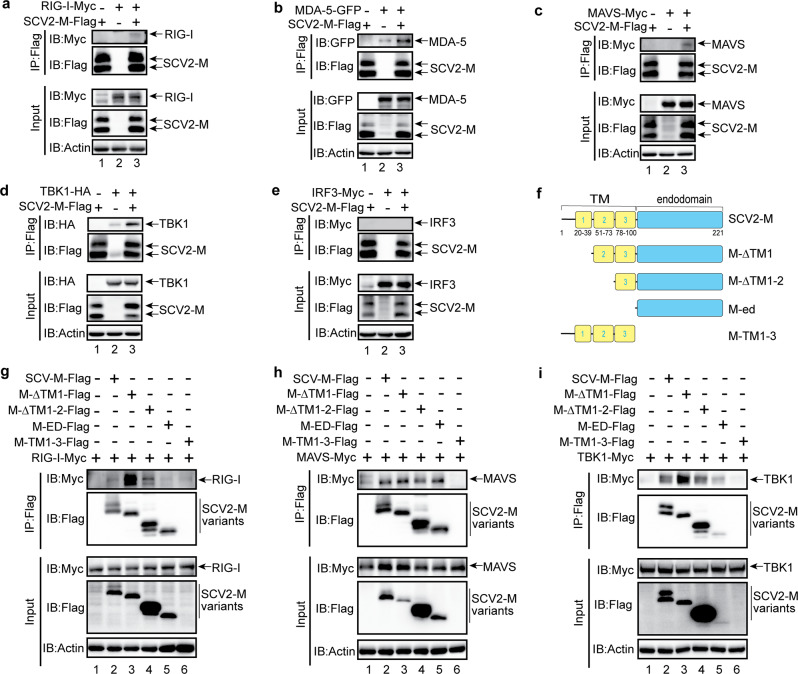
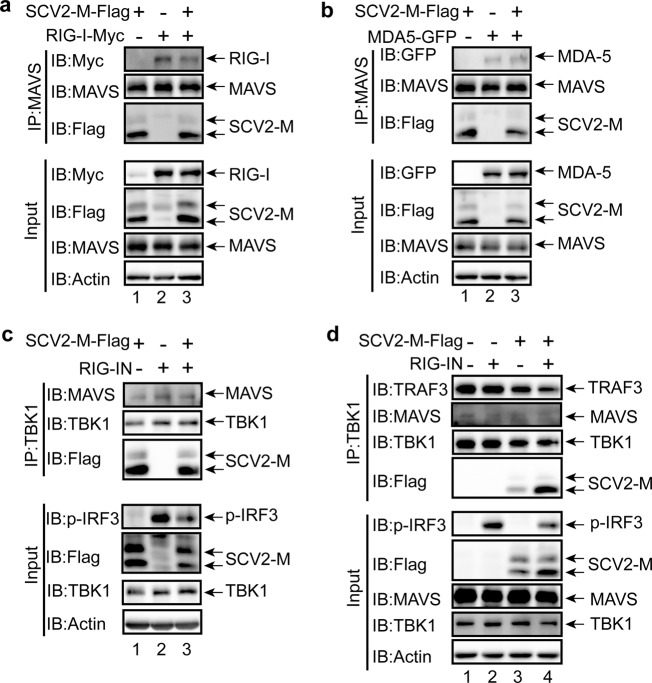
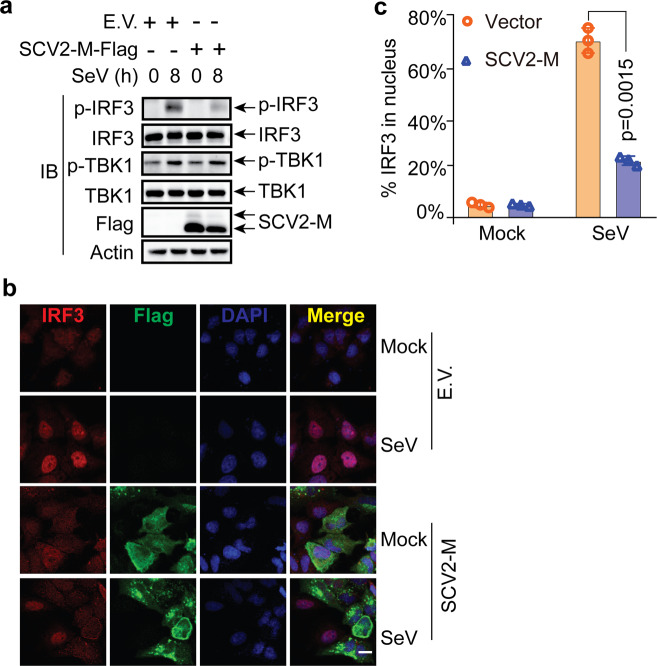
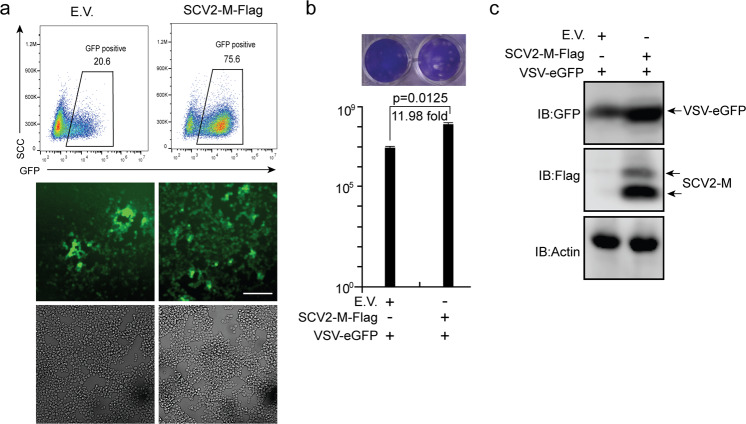
Coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has quickly spread worldwide and has affected more than 10 million individuals. A typical feature of COVID-19 is the suppression of type I and III interferon (IFN)-mediated antiviral immunity. However, the molecular mechanism by which SARS-CoV-2 evades antiviral immunity remains elusive. Here, we reported that the SARS-CoV-2 membrane (M) protein inhibits the production of type I and III IFNs induced by the cytosolic dsRNA-sensing pathway mediated by RIG-I/MDA-5-MAVS signaling. In addition, the SARS-CoV-2 M protein suppresses type I and III IFN induction stimulated by SeV infection or poly (I:C) transfection. Mechanistically, the SARS-CoV-2 M protein interacts with RIG-I, MAVS, and TBK1, thus preventing the formation of the multiprotein complex containing RIG-I, MAVS, TRAF3, and TBK1 and subsequently impeding the phosphorylation, nuclear translocation, and activation of IRF3. Consequently, ectopic expression of the SARS-CoV-2 M protein facilitates the replication of vesicular stomatitis virus. Taken together, these results indicate that the SARS-CoV-2 M protein antagonizes type I and III IFN production by targeting RIG-I/MDA-5 signaling, which subsequently attenuates antiviral immunity and enhances viral replication. This study provides insight into the interpretation of SARS-CoV-2-induced antiviral immune suppression and illuminates the pathogenic mechanism of COVID-19.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- SBK2020042706/Natural Science Foundation of Jiangsu Province (Jiangsu Provincial Natural Science Foundation)/International
- 2018M642662/China Postdoctoral Science Foundation/International
- 81901604/National Natural Science Foundation of China (National Science Foundation of China)/International
- 31730026/National Natural Science Foundation of China (National Science Foundation of China)/International
- 81525012/National Natural Science Foundation of China (National Science Foundation of China)/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
