

Hierarchical Multivalent Effects Control Influenza Host Specificity
- PMID: 33376792
- PMCID: PMC7760459
- DOI: 10.1021/acscentsci.0c01175
Hierarchical Multivalent Effects Control Influenza Host Specificity
Abstract
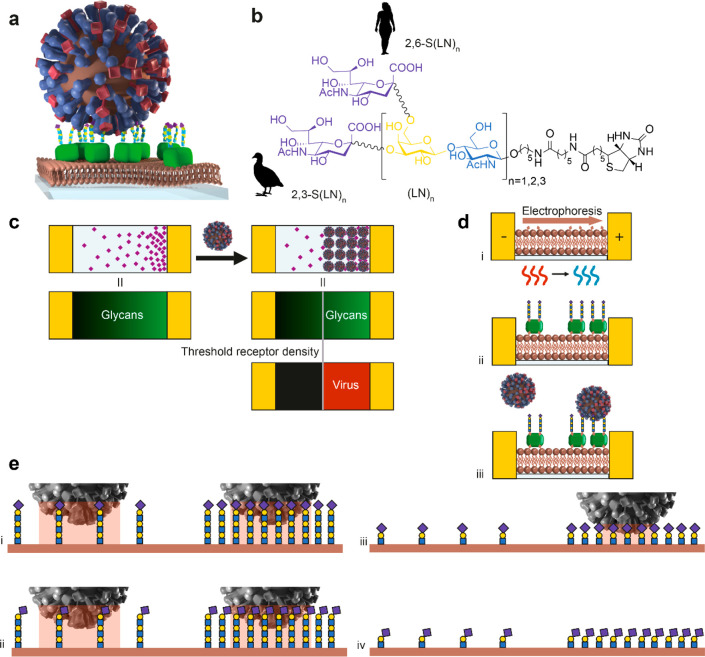
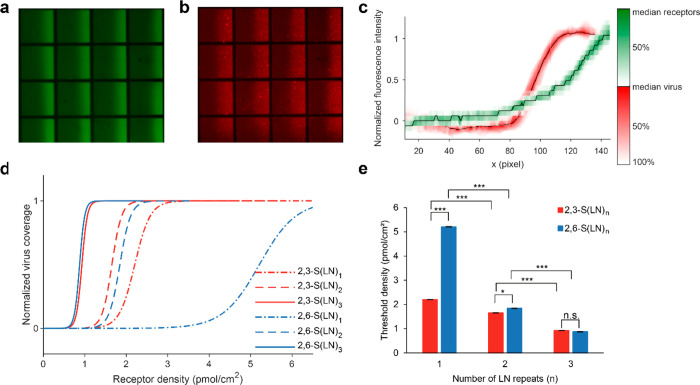
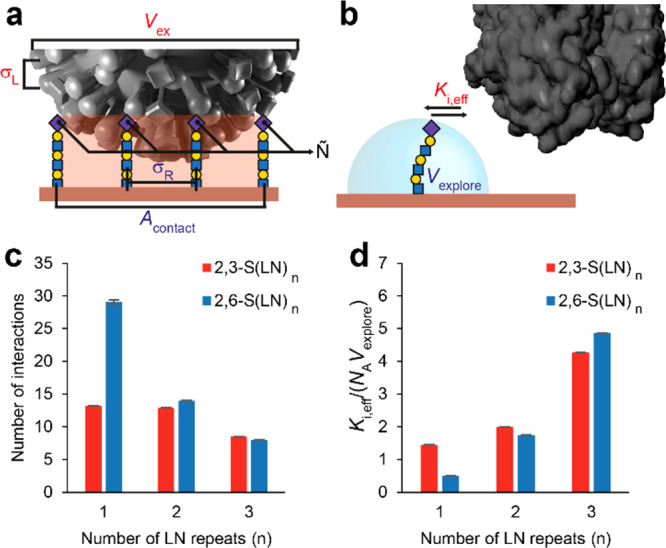
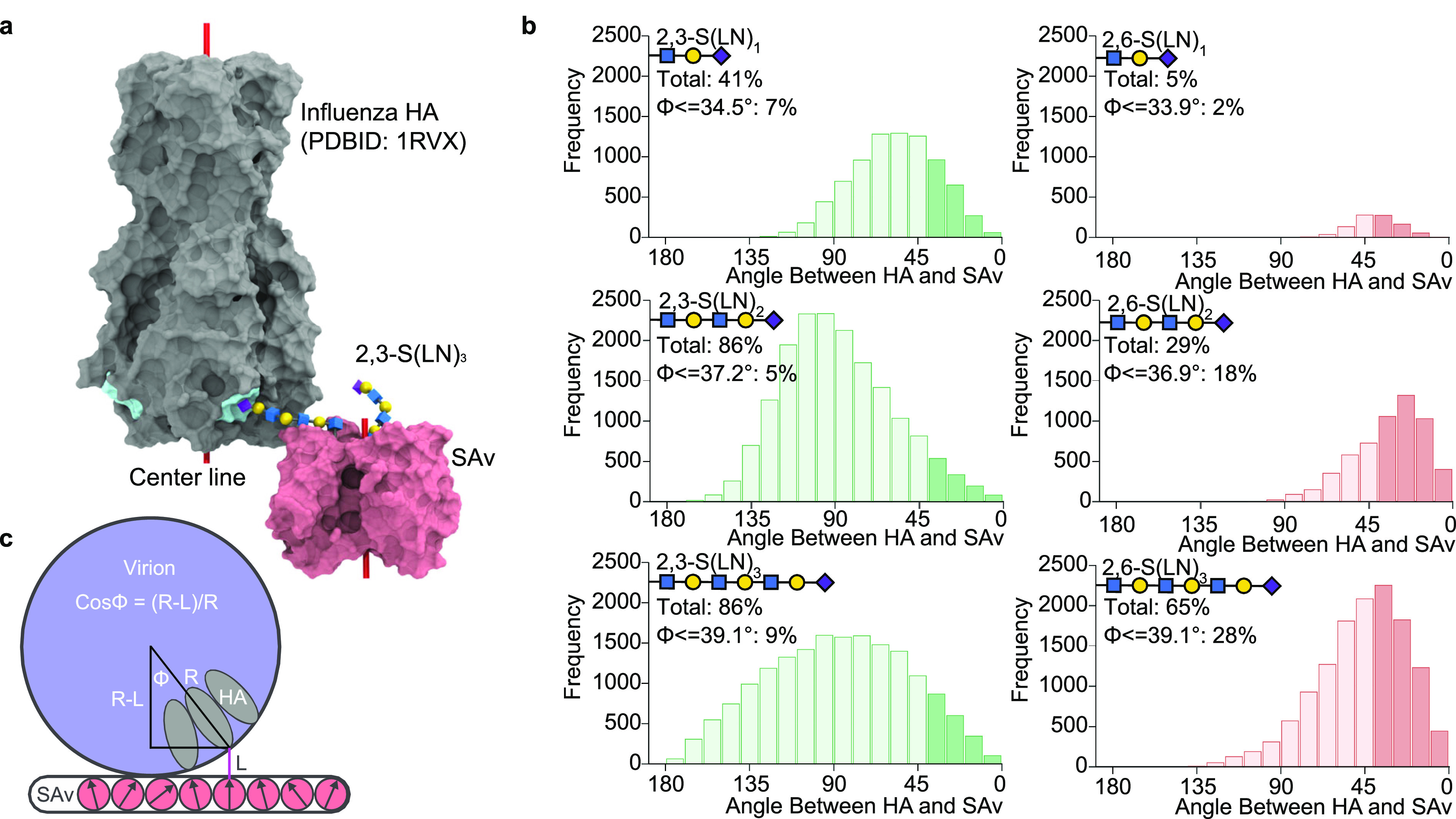
Understanding how emerging influenza viruses recognize host cells is critical in evaluating their zoonotic potential, pathogenicity, and transmissibility between humans. The surface of the influenza virus is covered with hemagglutinin (HA) proteins that can form multiple interactions with sialic acid-terminated glycans on the host cell surface. This multivalent binding affects the selectivity of the virus in ways that cannot be predicted from the individual receptor-ligand interactions alone. Here, we show that the intrinsic structural and energetic differences between the interactions of avian- or human-type receptors with influenza HA translate from individual site affinity and orientation through receptor length and density on the surface into virus avidity and specificity. We introduce a method to measure virus avidity using receptor density gradients. We found that influenza viruses attached stably to a surface at receptor densities that correspond to a minimum number of approximately 8 HA-glycan interactions, but more interactions were required if the receptors were short and human-type. Thus, the avidity and specificity of influenza viruses for a host cell depend not on the sialic acid linkage alone but on a combination of linkage and the length and density of receptors on the cell surface. Our findings suggest that threshold receptor densities play a key role in virus tropism, which is a predicting factor for both their virulence and zoonotic potential.
© 2020 American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Schaapveld M.; Aleman B. M. P.; van Eggermond A. M.; Janus C. P. M.; Krol A. D. G.; van der Maazen R. W. M.; Roesink J.; Raemaekers J. M. M.; de Boer J. P.; Zijlstra J. M.; et al. Emergence of a Novel Swine-Origin Influenza A (H1N1) Virus in Humans. N. Engl. J. Med. 2009, 360 (25), 2605–2615. 10.1056/NEJMoa0903810. - DOI - PubMed
-
- Maines T. R.; Chen L. M.; Van Hoeven N.; Tumpey T. M.; Blixt O.; Belser J. A.; Gustin K. M.; Pearce M. B.; Pappas C.; Stevens J.; et al. Effect of Receptor Binding Domain Mutations on Receptor Binding and Transmissibility of Avian Influenza H5N1 Viruses. Virology 2011, 413 (1), 139–147. 10.1016/j.virol.2011.02.015. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
