

The RNA phosphatase PIR-1 regulates endogenous small RNA pathways in C. elegans
- PMID: 33378643
- PMCID: PMC7867642
- DOI: 10.1016/j.molcel.2020.12.004
The RNA phosphatase PIR-1 regulates endogenous small RNA pathways in C. elegans
Abstract
Eukaryotic cells regulate 5'-triphosphorylated RNAs (ppp-RNAs) to promote cellular functions and prevent recognition by antiviral RNA sensors. For example, RNA capping enzymes possess triphosphatase domains that remove the γ phosphates of ppp-RNAs during RNA capping. Members of the closely related PIR-1 (phosphatase that interacts with RNA and ribonucleoprotein particle 1) family of RNA polyphosphatases remove both the β and γ phosphates from ppp-RNAs. Here, we show that C. elegans PIR-1 dephosphorylates ppp-RNAs made by cellular RNA-dependent RNA polymerases (RdRPs) and is required for the maturation of 26G-RNAs, Dicer-dependent small RNAs that regulate thousands of genes during spermatogenesis and embryogenesis. PIR-1 also regulates the CSR-1 22G-RNA pathway and has critical functions in both somatic and germline development. Our findings suggest that PIR-1 modulates both Dicer-dependent and Dicer-independent Argonaute pathways and provide insight into how cells and viruses use a conserved RNA phosphatase to regulate and respond to ppp-RNA species.
Keywords: RNA binding proteins; RNA phosphatase; RNAi; double-stranded RNAs; embryogenesis; germline gene regulation; germline small RNAs; mRNA regulation; regulation of triphosphorylated RNA; spermatogenesis.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



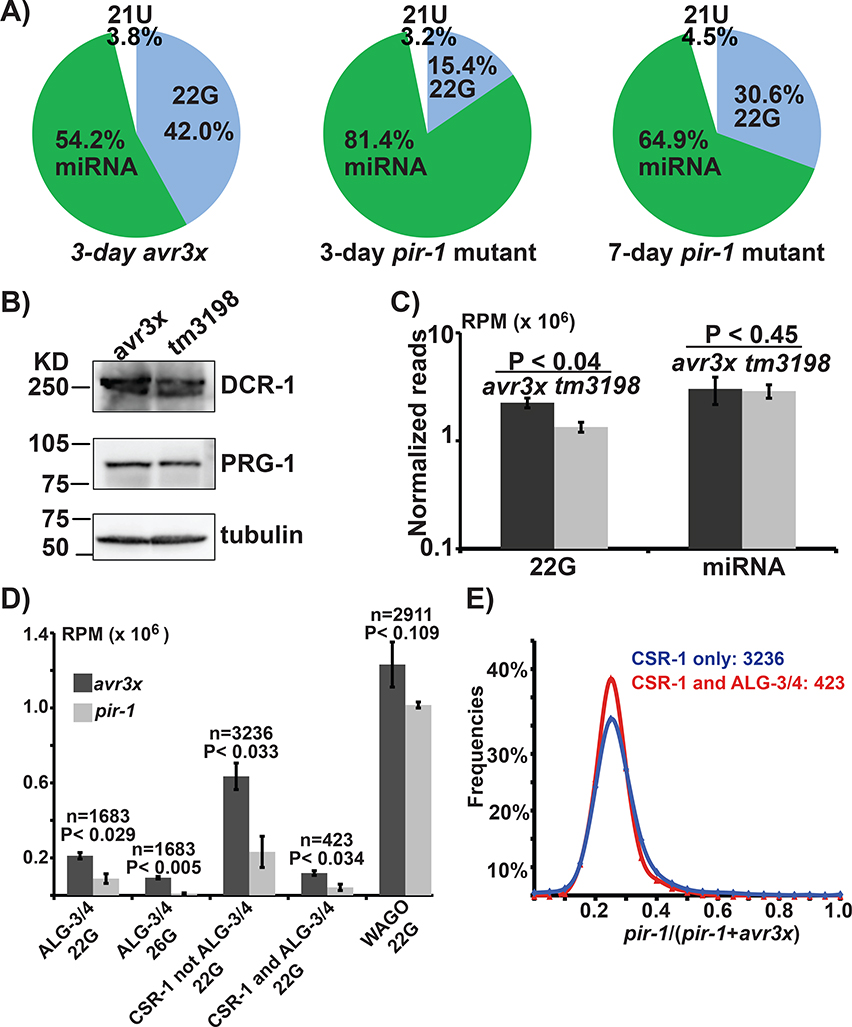
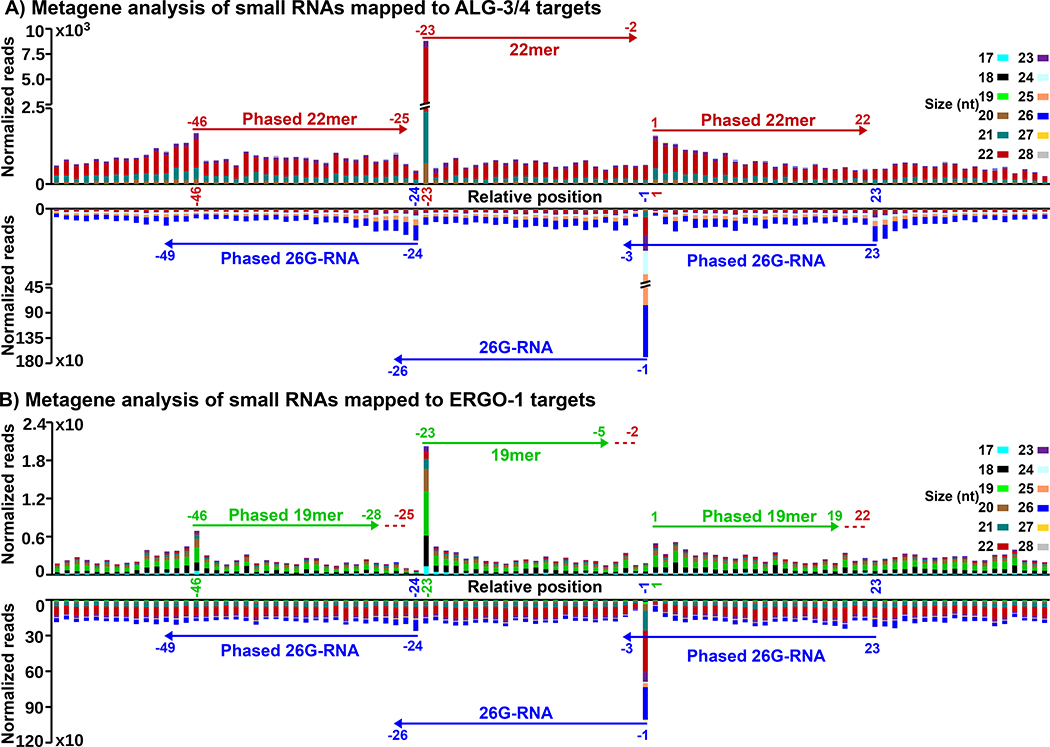

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
