

Dysregulation of sonic hedgehog signaling causes hearing loss in ciliopathy mouse models
- PMID: 33382037
- PMCID: PMC7806262
- DOI: 10.7554/eLife.56551
Dysregulation of sonic hedgehog signaling causes hearing loss in ciliopathy mouse models
Abstract
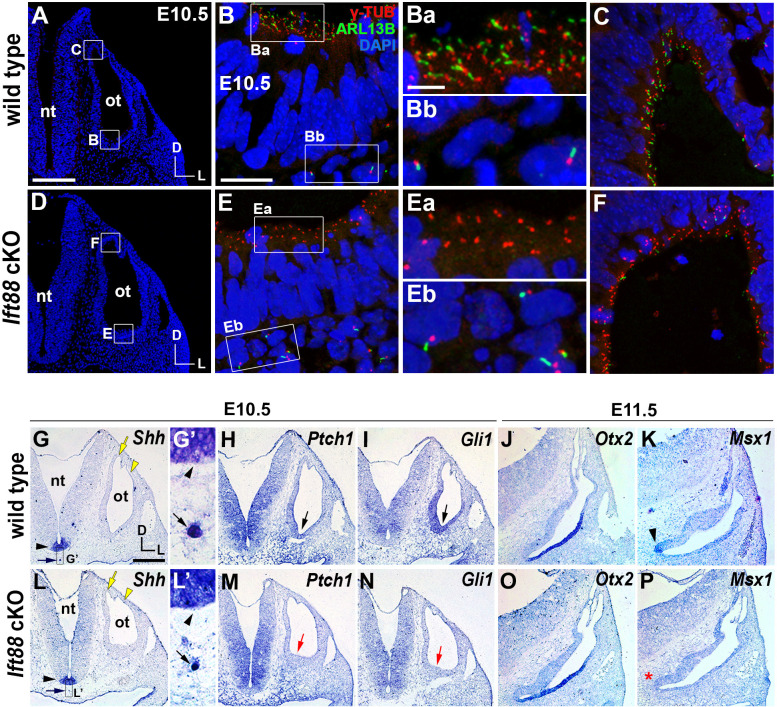
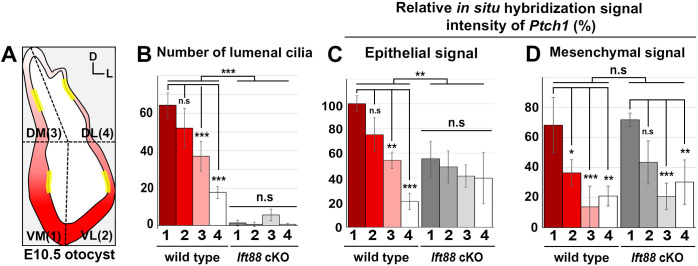
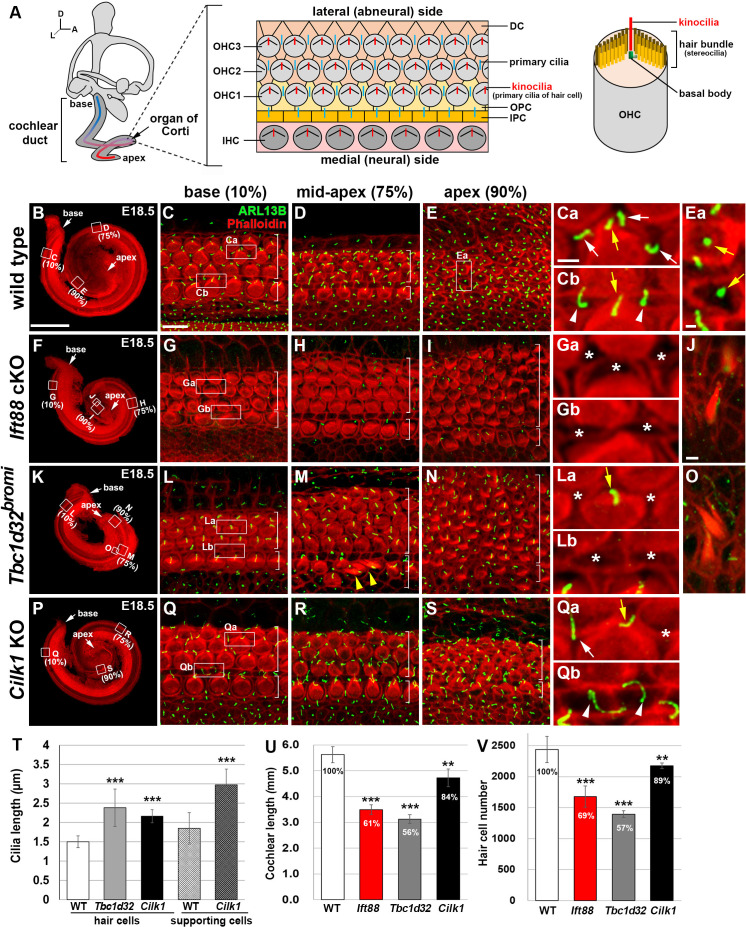
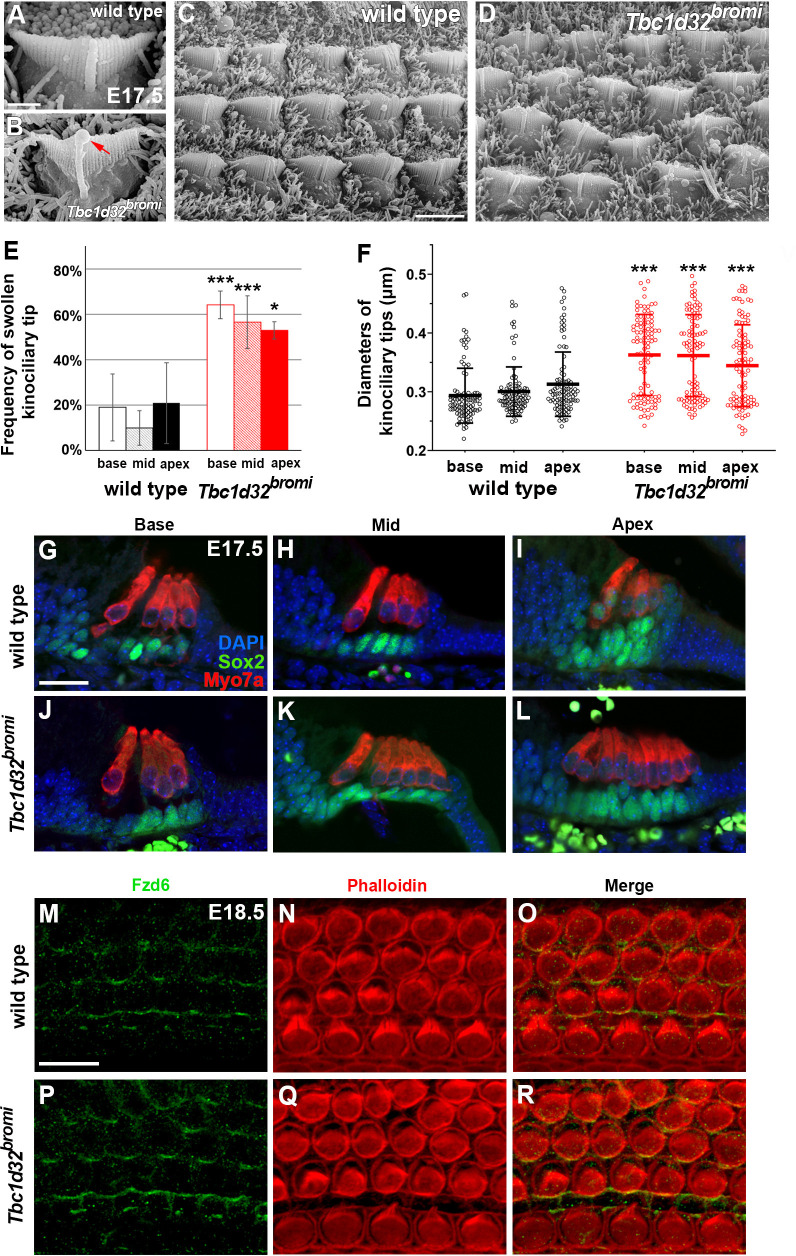
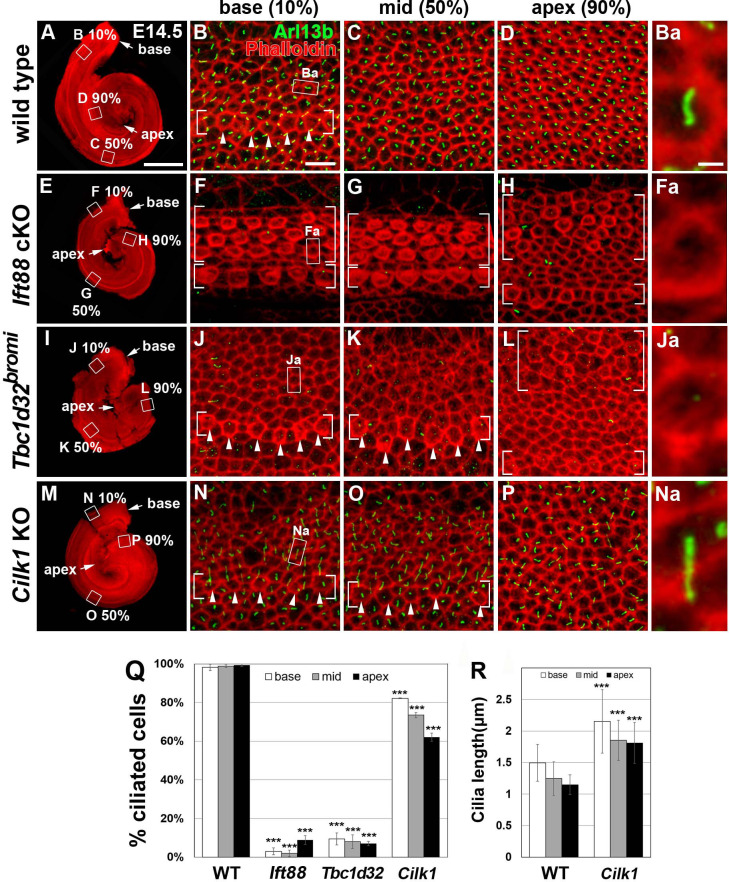
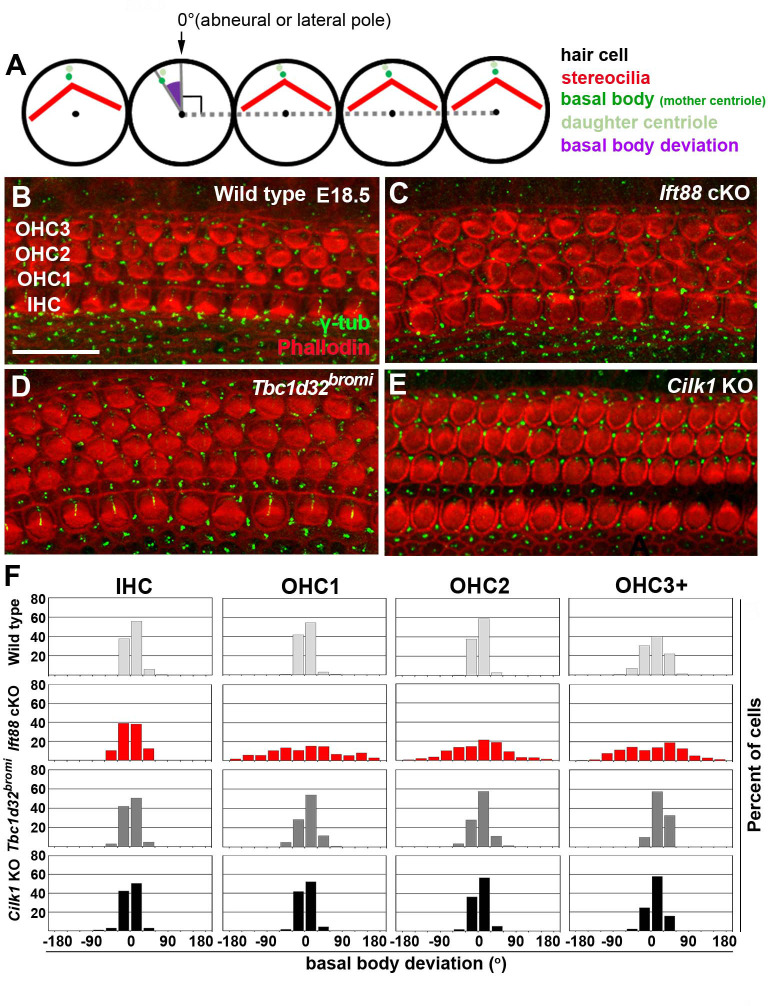
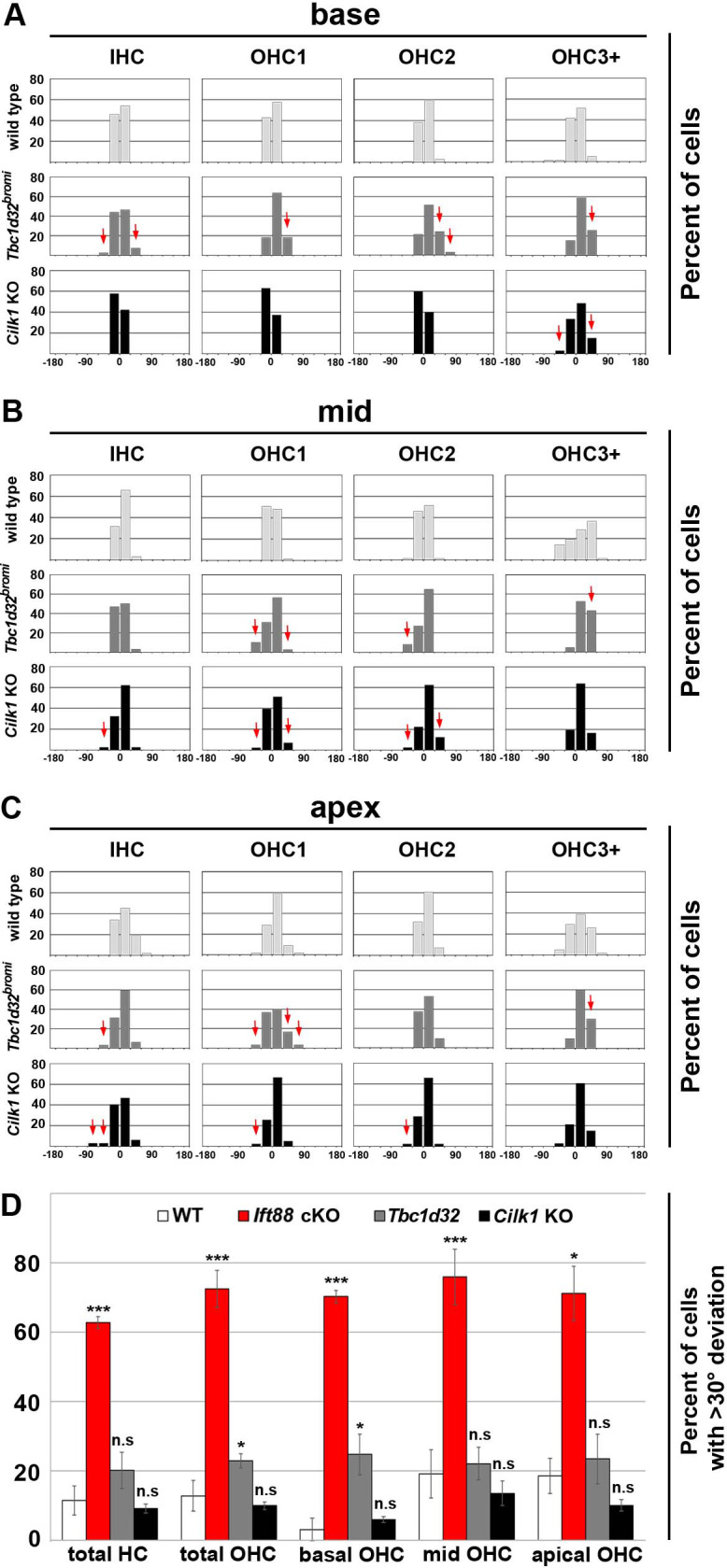
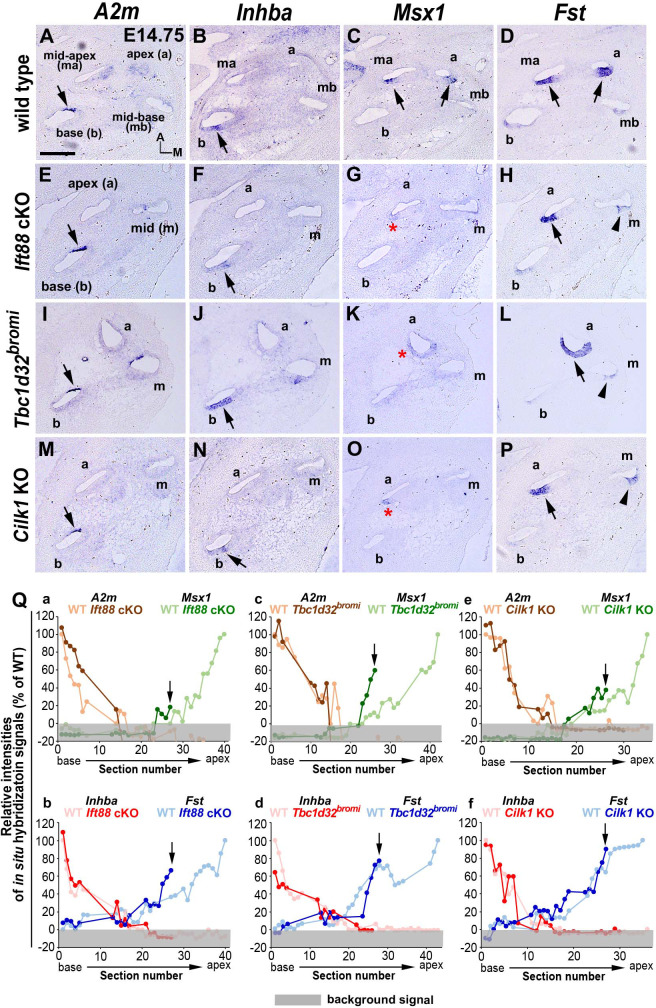
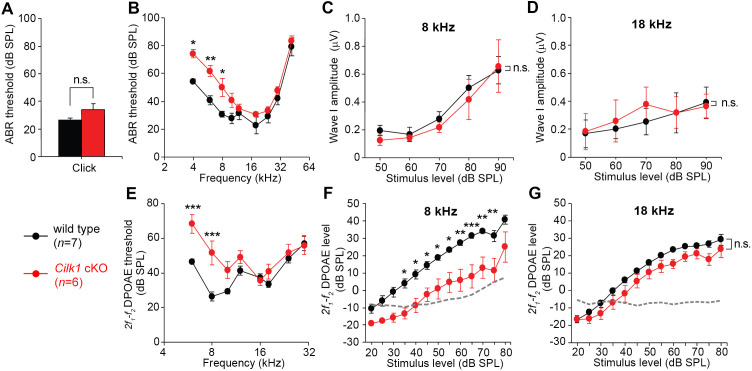
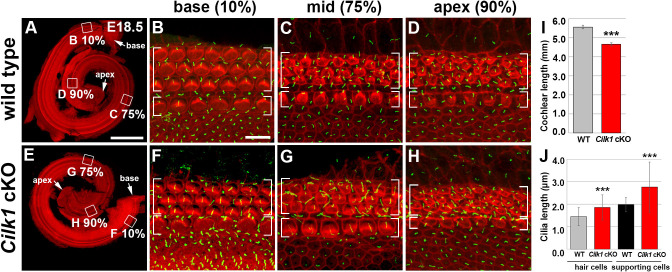
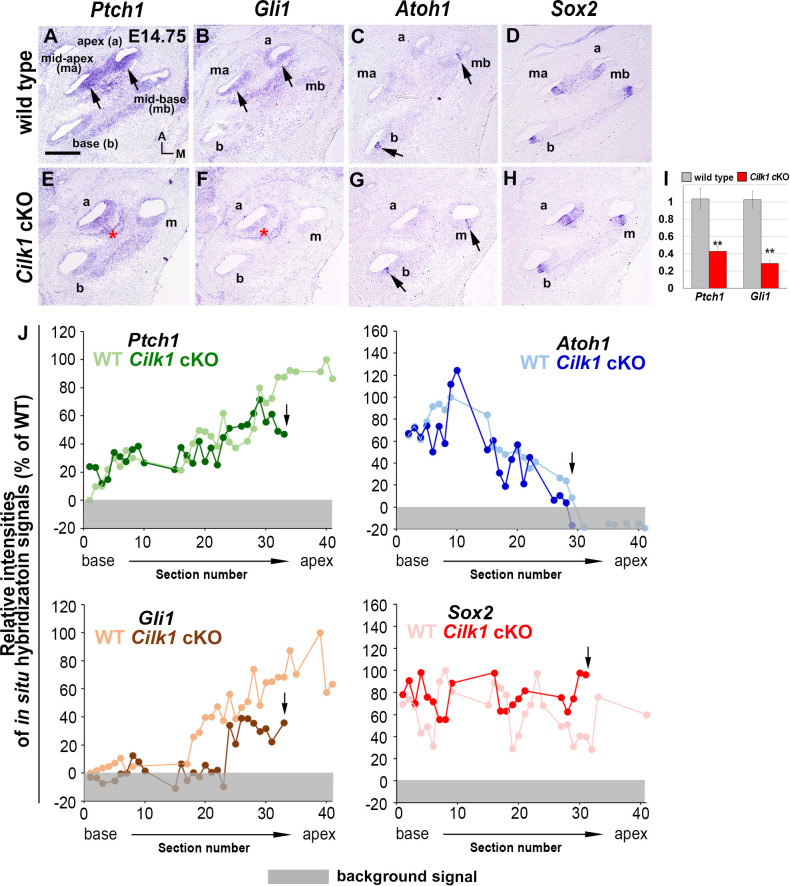
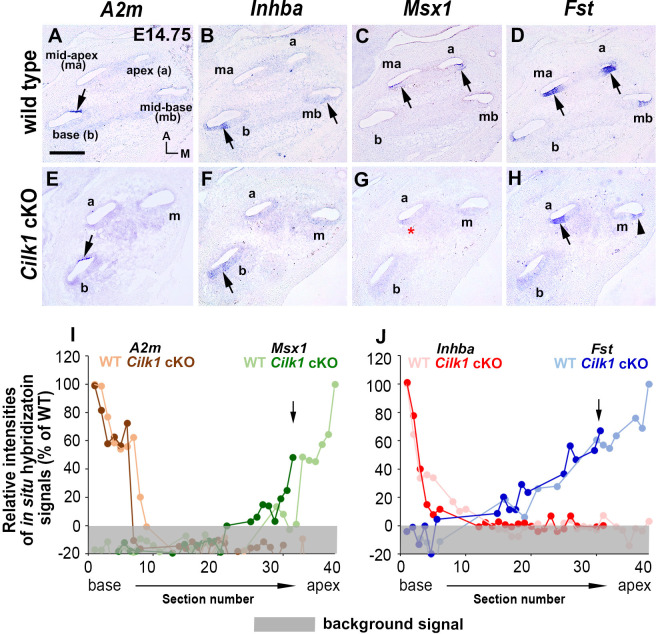
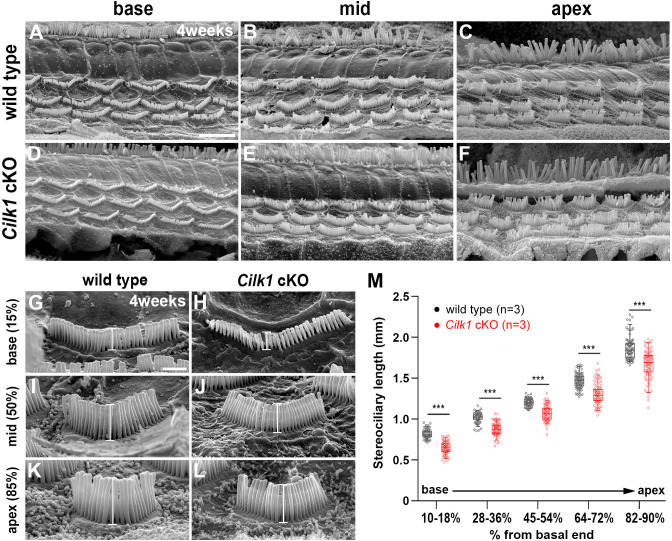
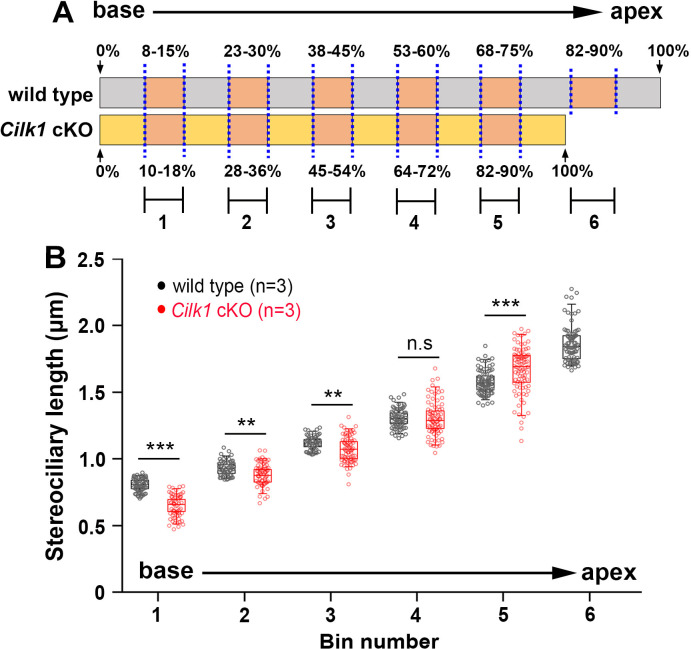
Defective primary cilia cause a range of diseases known as ciliopathies, including hearing loss. The etiology of hearing loss in ciliopathies, however, remains unclear. We analyzed cochleae from three ciliopathy mouse models exhibiting different ciliogenesis defects: Intraflagellar transport 88 (Ift88), Tbc1d32 (a.k.a. bromi), and Cilk1 (a.k.a. Ick) mutants. These mutants showed multiple developmental defects including shortened cochlear duct and abnormal apical patterning of the organ of Corti. Although ciliogenic defects in cochlear hair cells such as misalignment of the kinocilium are often associated with the planar cell polarity pathway, our results showed that inner ear defects in these mutants are primarily due to loss of sonic hedgehog signaling. Furthermore, an inner ear-specific deletion of Cilk1 elicits low-frequency hearing loss attributable to cellular changes in apical cochlear identity that is dedicated to low-frequency sound detection. This type of hearing loss may account for hearing deficits in some patients with ciliopathies.
Keywords: ciliopathies; developmental biology; hearing loss; mouse; primary cilia; sonic hedgehog.
© 2020, Moon et al.
Conflict of interest statement
KM, JM, HM, HK, HK, HK, JB No competing interests declared
Figures
References
-
- Abdelhamed ZA, Natarajan S, Wheway G, Inglehearn CF, Toomes C, Johnson CA, Jagger DJ. The Meckel-Gruber syndrome protein TMEM67 controls basal body positioning and epithelial branching morphogenesis in mice via the non-canonical wnt pathway. Disease Models & Mechanisms. 2015;8:527–541. doi: 10.1242/dmm.019083. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
