

Bulges control pri-miRNA processing in a position and strand-dependent manner
- PMID: 33382955
- PMCID: PMC8582997
- DOI: 10.1080/15476286.2020.1868139
Bulges control pri-miRNA processing in a position and strand-dependent manner
Abstract
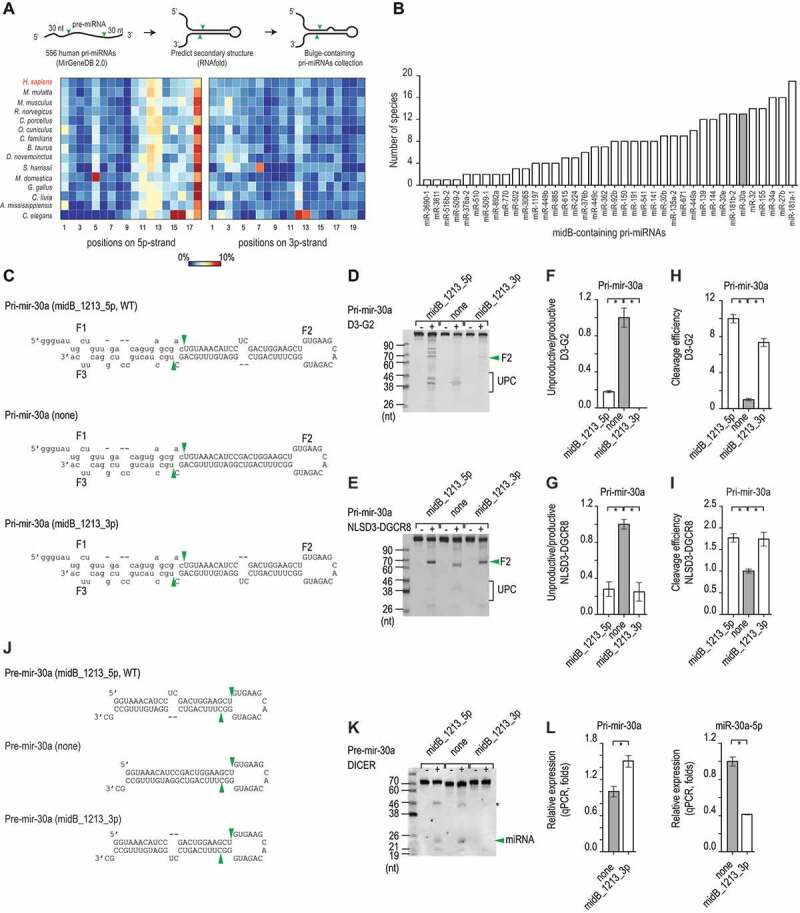
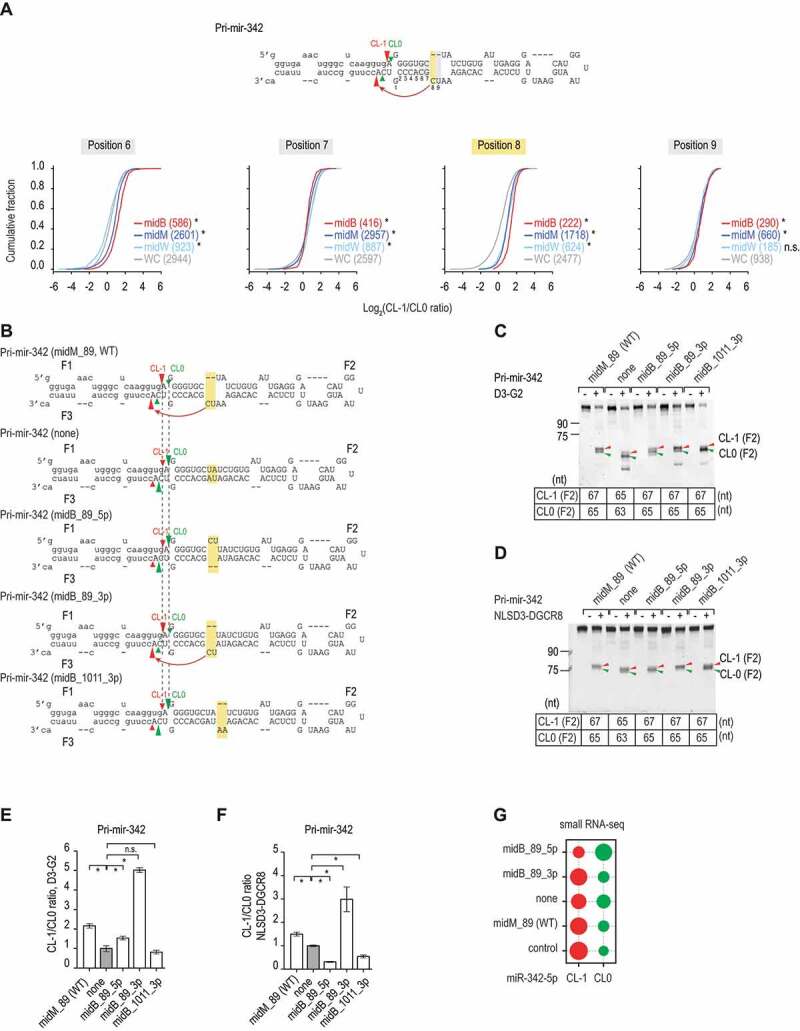
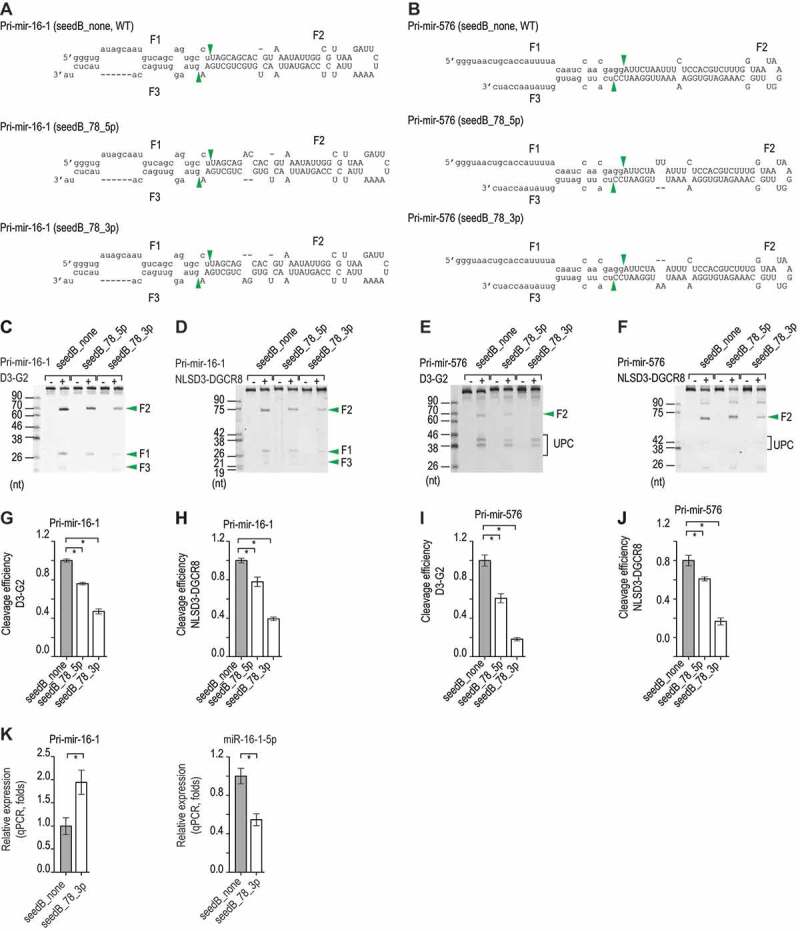
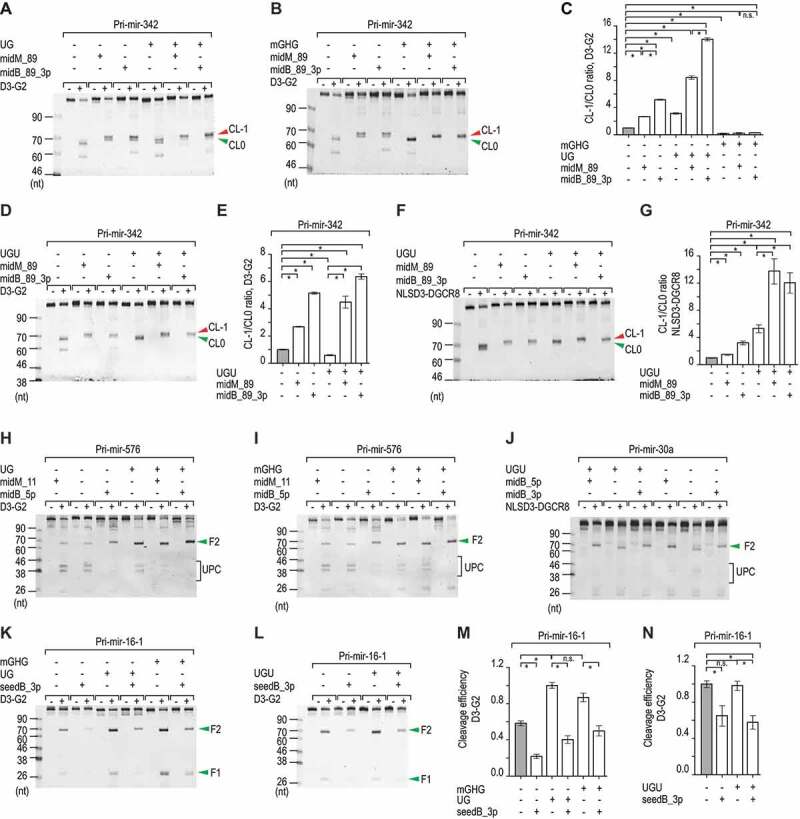
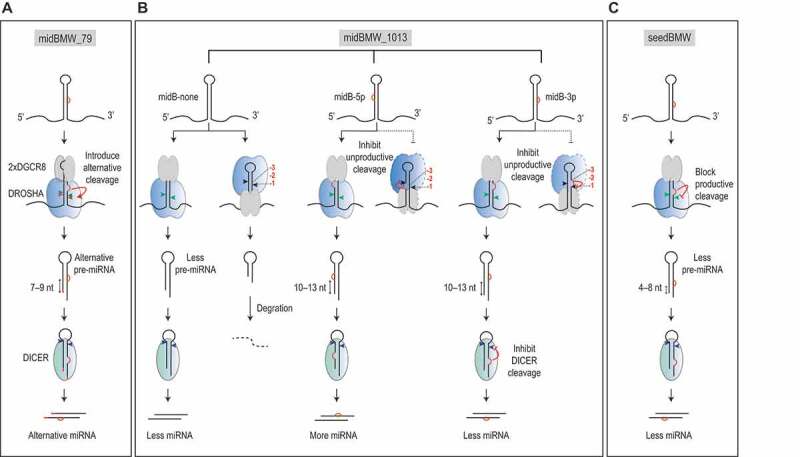
MicroRNAs (miRNAs) play critical roles in gene expression and numerous human diseases. The success of miRNA biogenesis is largely determined by the primary miRNA (pri-miRNA) processing by the DROSHA-DGCR8 complex, called Microprocessor. Here, we analysed the high-throughput pri-miRNA processing assays and secondary structures of pri-miRNAs to investigate the roles of bulges in the pri-miRNA processing. We found that bulges in multiple places control both the cleavage efficiency and accuracy of pri-miRNA processing. These bulges were shown to act on Microprocessor via its catalytic subunit, DROSHA, and function in a position and strand-dependent manner. Interestingly, we discovered that the enriched and conserved bulges, called midB, can correct DROSHA orientation on pri-miRNAs, thereby enhancing production of miRNAs. The revealed functions of the bulges help improve our understanding of pri-miRNA processing and suggest their potential roles in miRNA biogenesis regulation.
Keywords: Bulges; DGCR8; DROSHA; Microprocessor; miRNA biogenesis.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
Similar articles
-
A central role for the primary microRNA stem in guiding the position and efficiency of Drosha processing of a viral pri-miRNA.RNA. 2014 Jul;20(7):1068-77. doi: 10.1261/rna.044537.114. Epub 2014 May 22. RNA. 2014. PMID: 24854622 Free PMC article.
-
Molecular Basis for the Single-Nucleotide Precision of Primary microRNA Processing.Mol Cell. 2019 Feb 7;73(3):505-518.e5. doi: 10.1016/j.molcel.2018.11.005. Epub 2018 Dec 13. Mol Cell. 2019. PMID: 30554947
-
Structural Basis for pri-miRNA Recognition by Drosha.Mol Cell. 2020 May 7;78(3):423-433.e5. doi: 10.1016/j.molcel.2020.02.024. Epub 2020 Mar 27. Mol Cell. 2020. PMID: 32220645
-
Orientation of Human Microprocessor on Primary MicroRNAs.Biochemistry. 2019 Jan 29;58(4):189-198. doi: 10.1021/acs.biochem.8b00944. Epub 2018 Dec 10. Biochemistry. 2019. PMID: 30481000 Review.
-
MicroRNA biogenesis: isolation and characterization of the microprocessor complex.Methods Mol Biol. 2006;342:33-47. doi: 10.1385/1-59745-123-1:33. Methods Mol Biol. 2006. PMID: 16957365 Review.
Cited by
-
Secondary structure RNA elements control the cleavage activity of DICER.Nat Commun. 2022 Apr 19;13(1):2138. doi: 10.1038/s41467-022-29822-3. Nat Commun. 2022. PMID: 35440644 Free PMC article.
-
Consequences of genetic variants in miRNA genes.Comput Struct Biotechnol J. 2022 Nov 19;20:6443-6457. doi: 10.1016/j.csbj.2022.11.036. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36467588 Free PMC article. Review.
-
Assessing Microprocessor complex mutations with a Microsensor system.RNA. 2025 Jun 16;31(7):896-915. doi: 10.1261/rna.080338.124. RNA. 2025. PMID: 40328469
-
TIM proteins and microRNAs: distinct impact and promising interactions on transplantation immunity.Front Immunol. 2024 Nov 22;15:1500228. doi: 10.3389/fimmu.2024.1500228. eCollection 2024. Front Immunol. 2024. PMID: 39650660 Free PMC article. Review.
-
The conserved single-cleavage mechanism of animal DROSHA enzymes.Commun Biol. 2021 Nov 25;4(1):1332. doi: 10.1038/s42003-021-02860-1. Commun Biol. 2021. PMID: 34824450 Free PMC article.
References
-
- Iwasaki S, Tomari Y.. Argonaute-mediated translational repression (and activation). Fly (Austin). 2009;3:205–208. - PubMed
-
- Jonas S, Izaurralde E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat Rev Genet. 2015;16:421–433. - PubMed
-
- Kawamata T, Making TY. RISC. Trends Biochem Sci. 2010;35:368–376. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases