

A circuit logic for sexually shared and dimorphic aggressive behaviors in Drosophila
- PMID: 33382967
- PMCID: PMC7856078
- DOI: 10.1016/j.cell.2020.11.048
A circuit logic for sexually shared and dimorphic aggressive behaviors in Drosophila
Erratum in
-
A circuit logic for sexually shared and dimorphic aggressive behaviors in Drosophila.Cell. 2021 Feb 4;184(3):847. doi: 10.1016/j.cell.2021.01.021. Cell. 2021. PMID: 33545040 Free PMC article. No abstract available.
Abstract
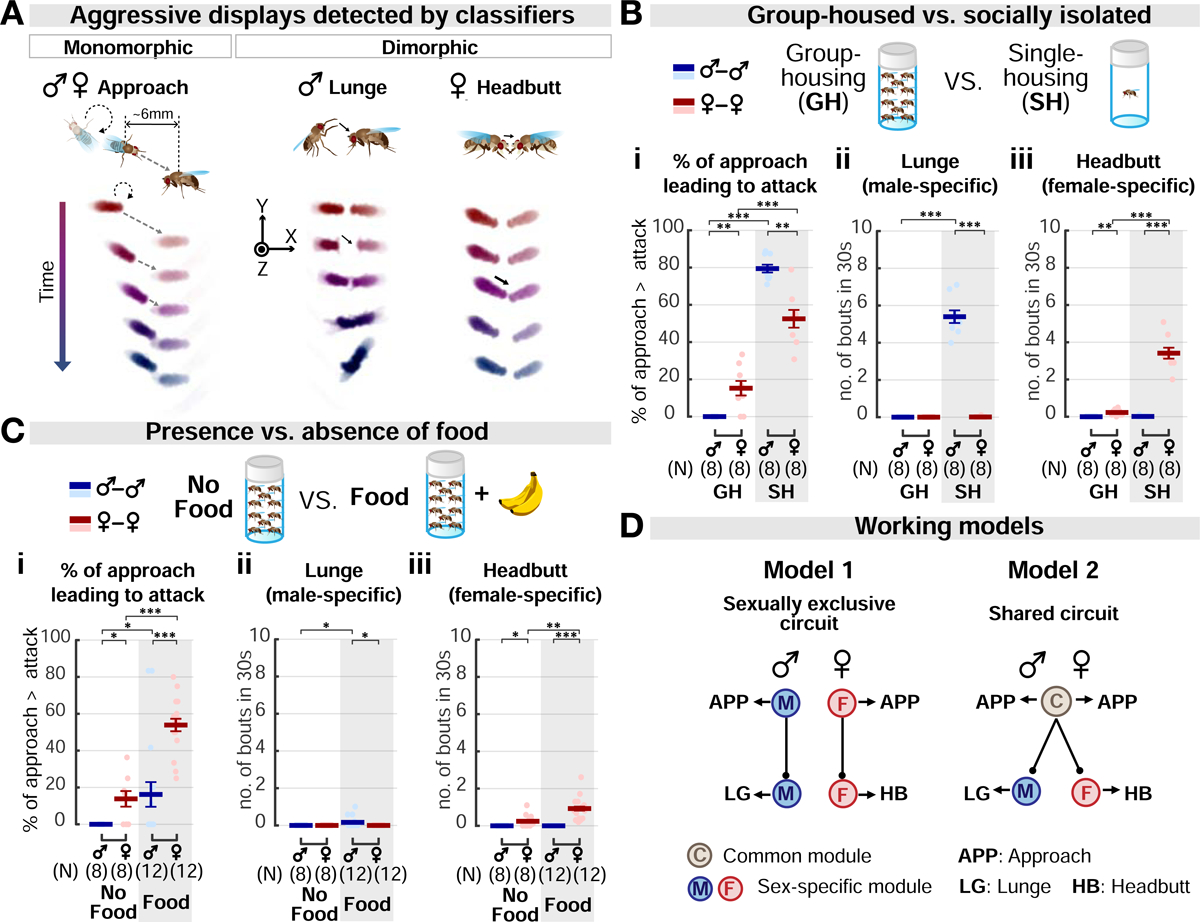
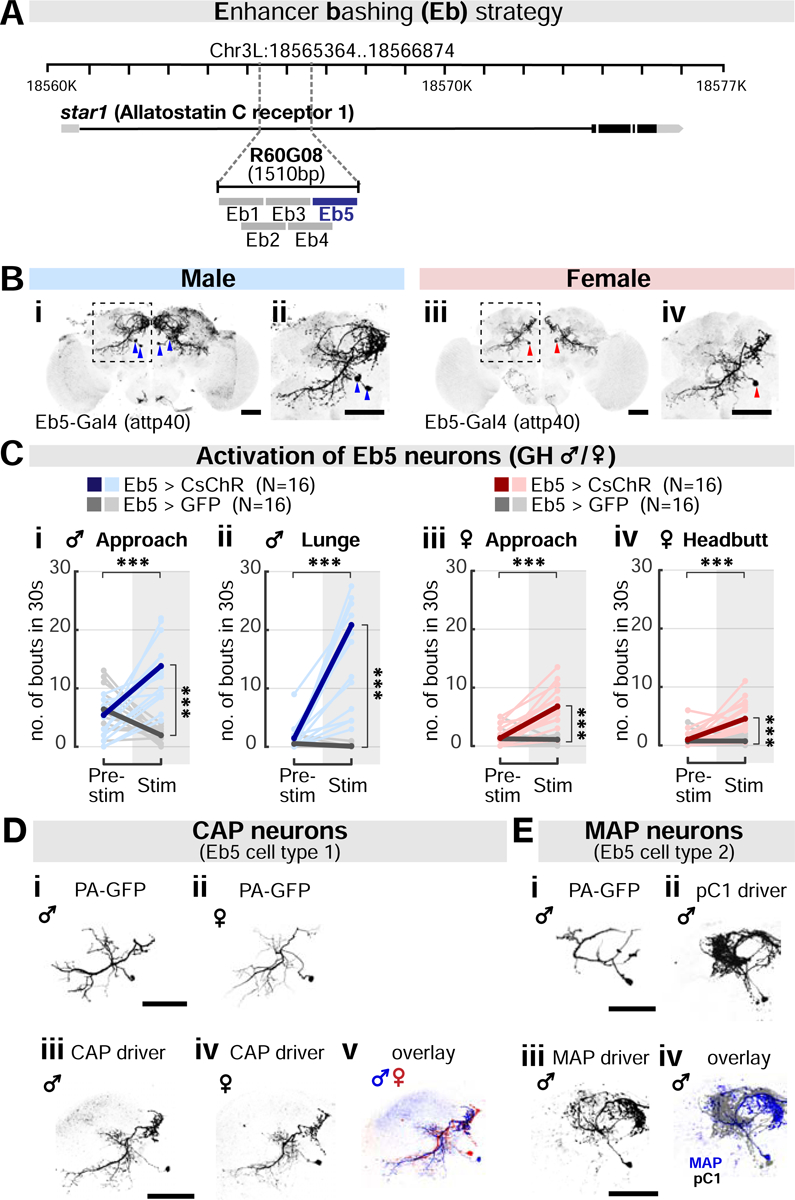
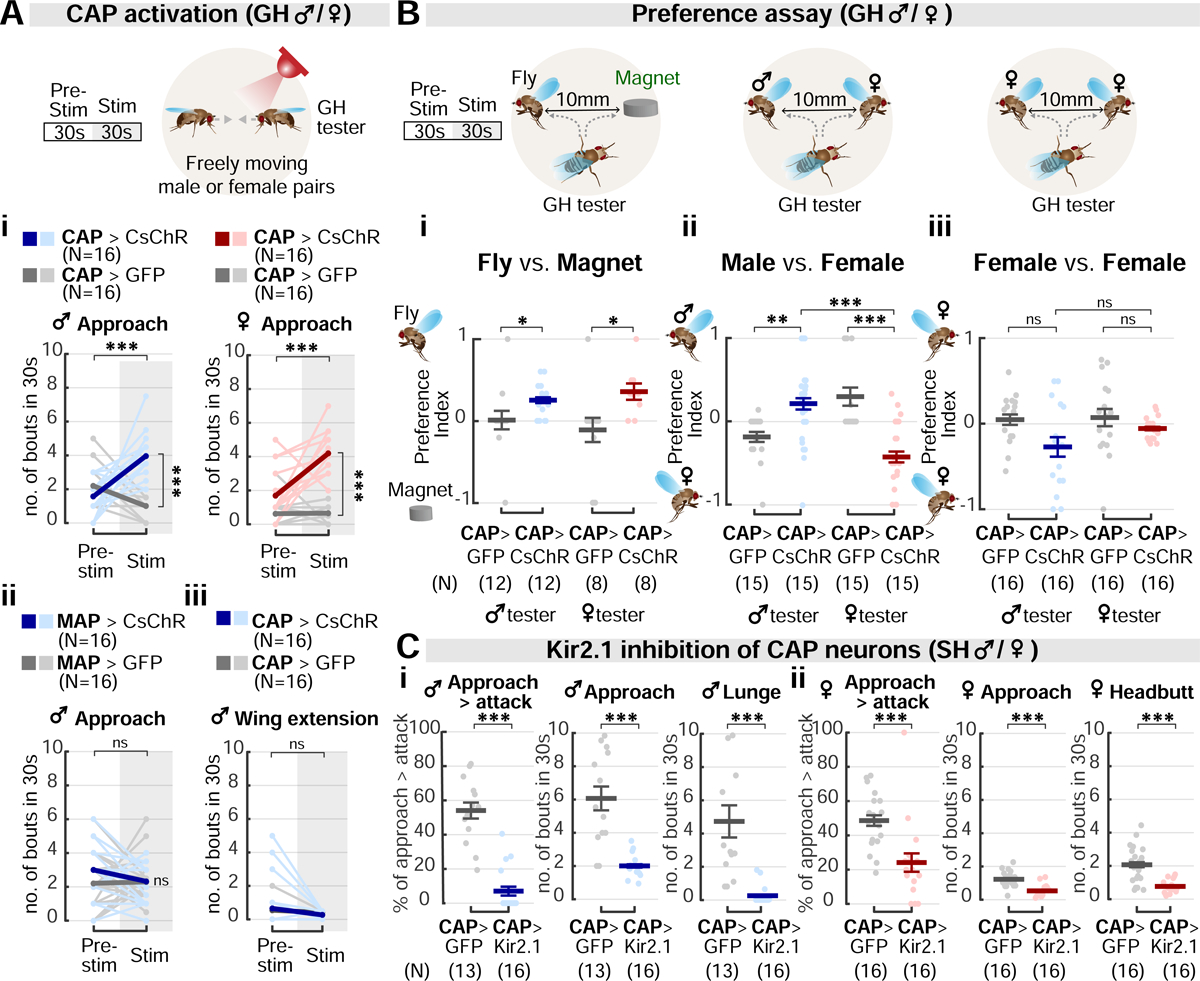
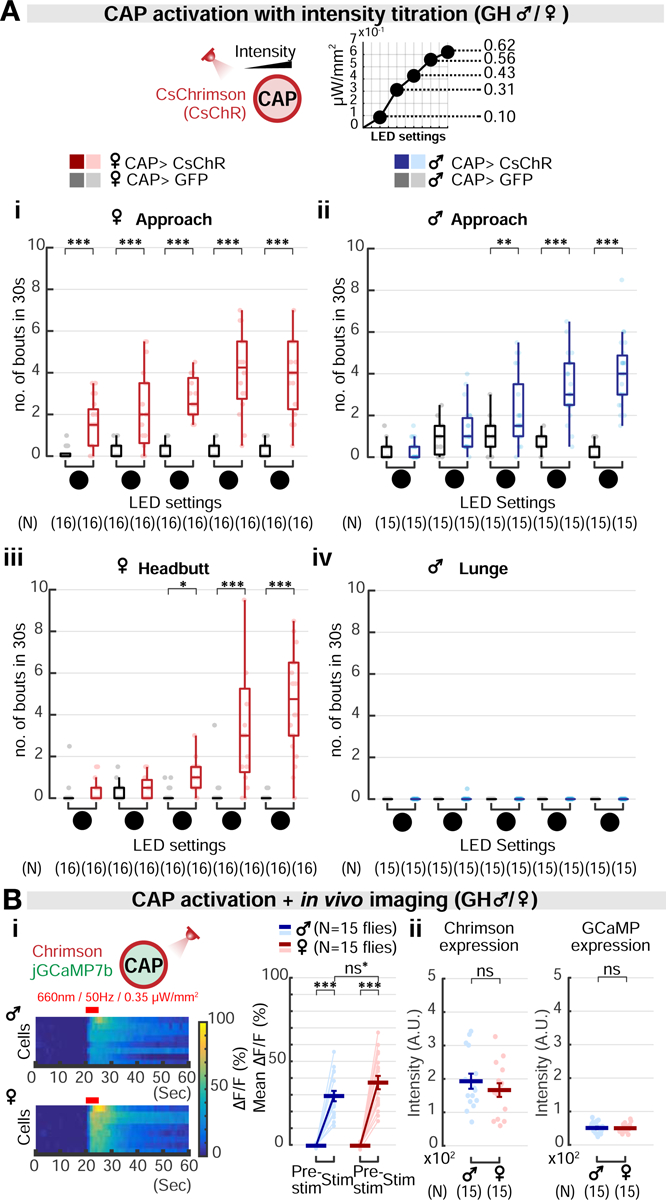
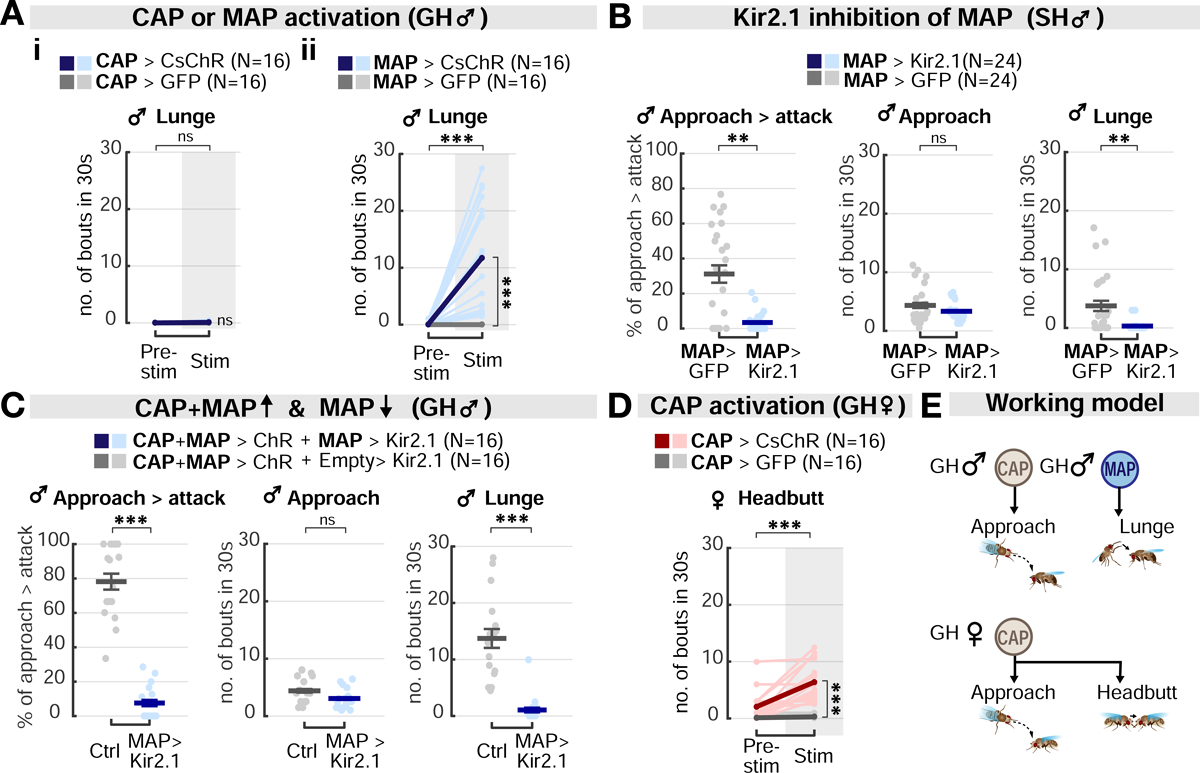
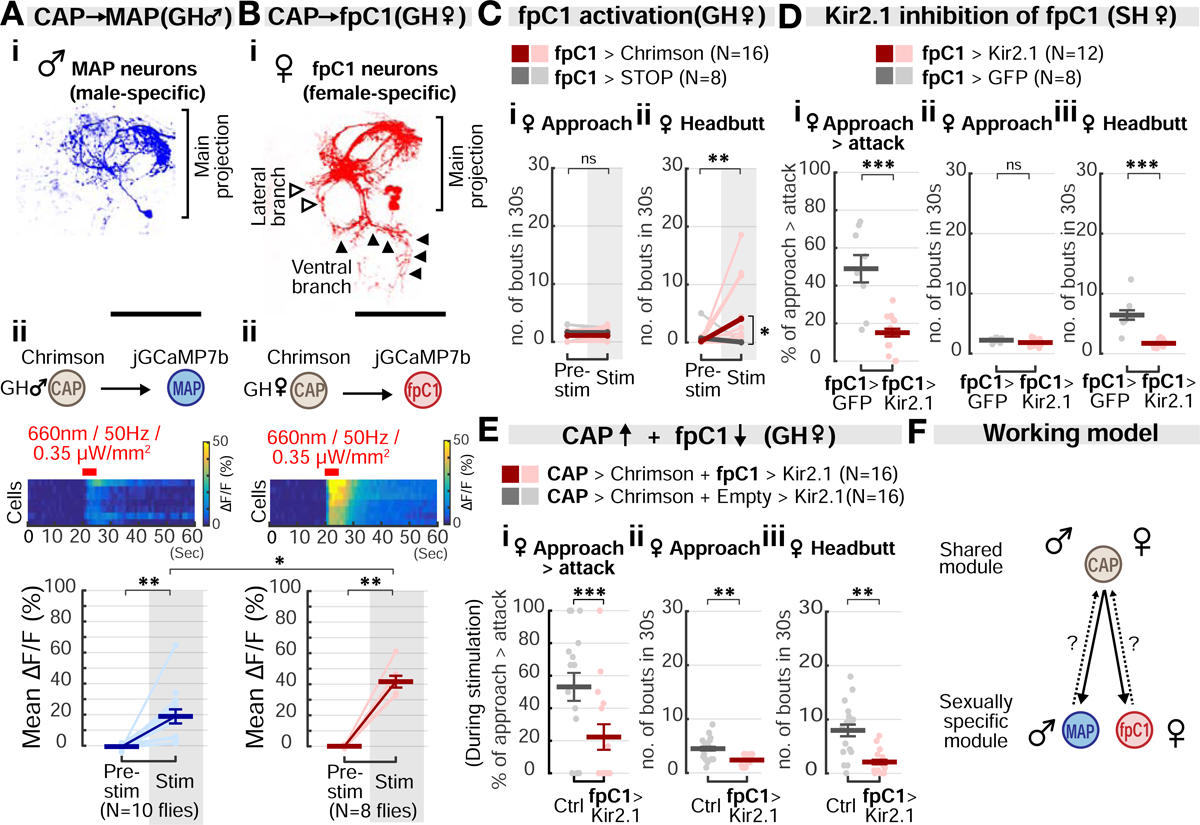
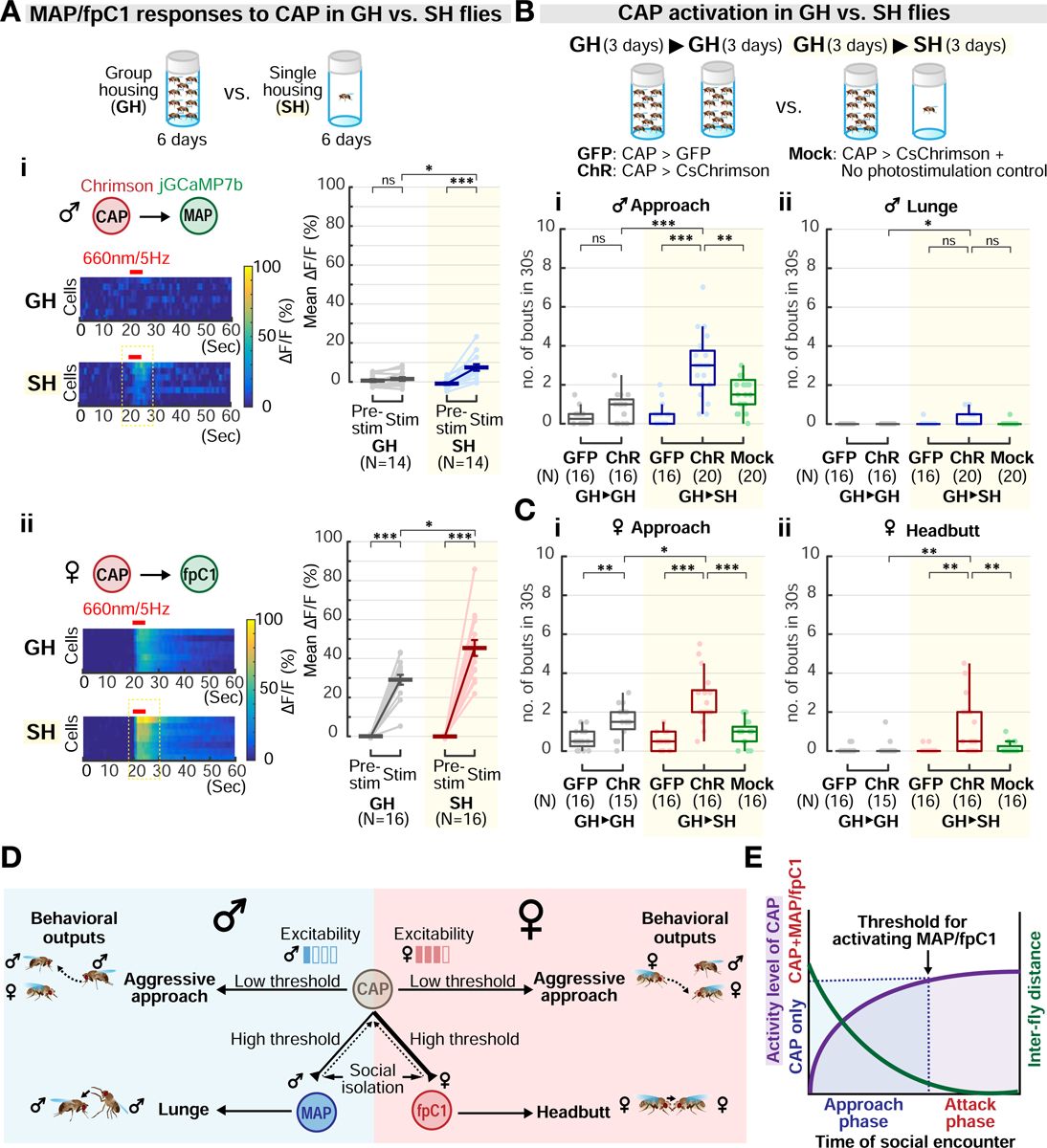
Aggression involves both sexually monomorphic and dimorphic actions. How the brain implements these two types of actions is poorly understood. We have identified three cell types that regulate aggression in Drosophila: one type is sexually shared, and the other two are sex specific. Shared common aggression-promoting (CAP) neurons mediate aggressive approach in both sexes, whereas functionally downstream dimorphic but homologous cell types, called male-specific aggression-promoting (MAP) neurons in males and fpC1 in females, control dimorphic attack. These symmetric circuits underlie the divergence of male and female aggressive behaviors, from their monomorphic appetitive/motivational to their dimorphic consummatory phases. The strength of the monomorphic → dimorphic functional connection is increased by social isolation in both sexes, suggesting that it may be a locus for isolation-dependent enhancement of aggression. Together, these findings reveal a circuit logic for the neural control of behaviors that include both sexually monomorphic and dimorphic actions, which may generalize to other organisms.
Keywords: Drosophila; appetitive behavior; consummatory behavior; sexual dimorphism.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Fly fighters.Nat Rev Neurosci. 2021 Mar;22(3):134-135. doi: 10.1038/s41583-021-00430-2. Nat Rev Neurosci. 2021. PMID: 33473202 No abstract available.
References
-
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Amanatides PG, Scherer SE, Li PW, Hoskins RA, Galle RF, et al. (2000). The genome sequence of Drosophila melanogaster. Science 287, 2185–2195. - PubMed
-
- Anderson DJ (2016). Circuit modules linking internal states and social behaviour in flies and mice. Nat. Rev. Neurosci 17, 692–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
