

Chromosome Segregation and Peptidoglycan Remodeling Are Coordinated at a Highly Stabilized Septal Pore to Maintain Bacterial Spore Development
- PMID: 33383000
- PMCID: PMC8048138
- DOI: 10.1016/j.devcel.2020.12.006
Chromosome Segregation and Peptidoglycan Remodeling Are Coordinated at a Highly Stabilized Septal Pore to Maintain Bacterial Spore Development
Abstract
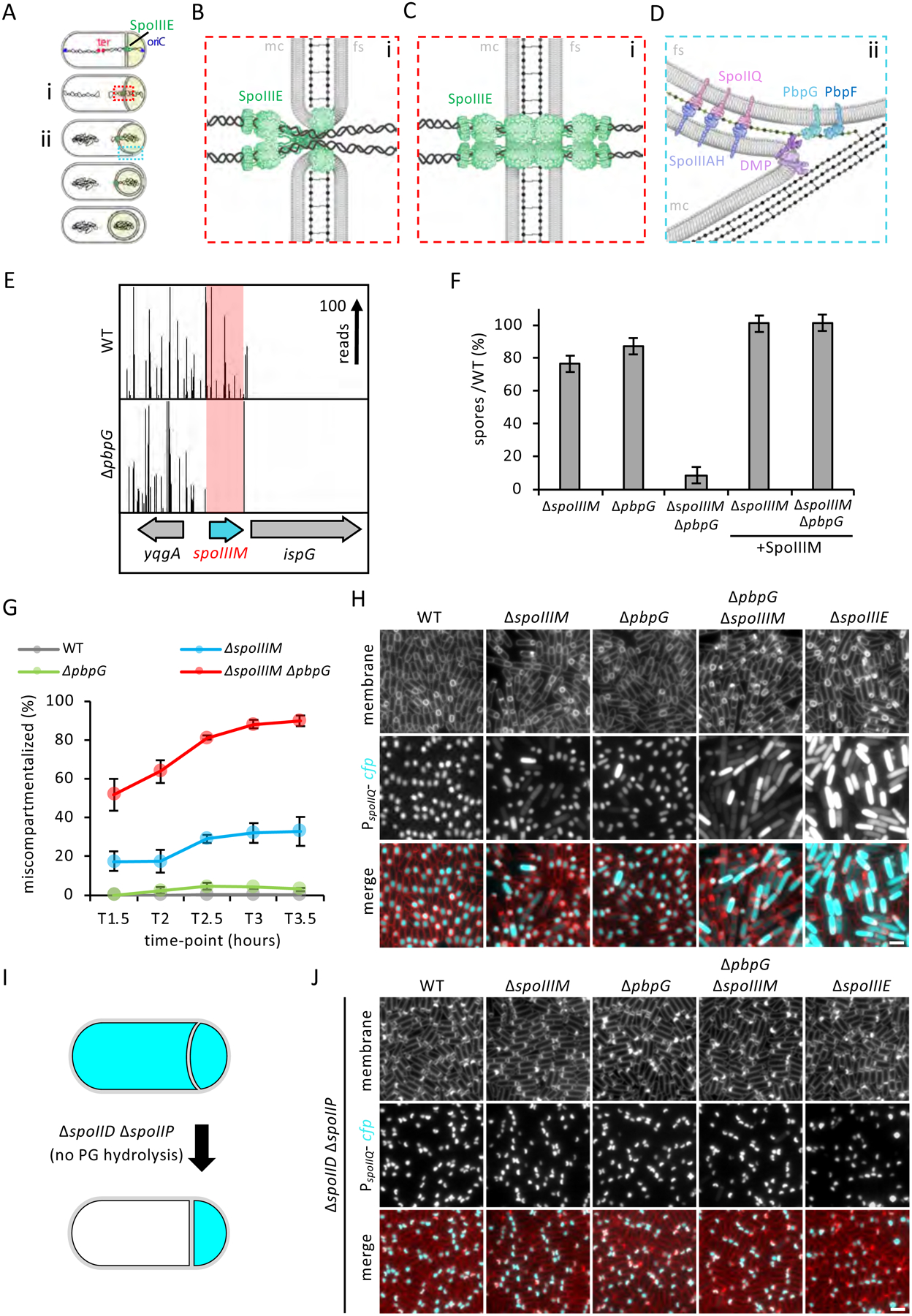
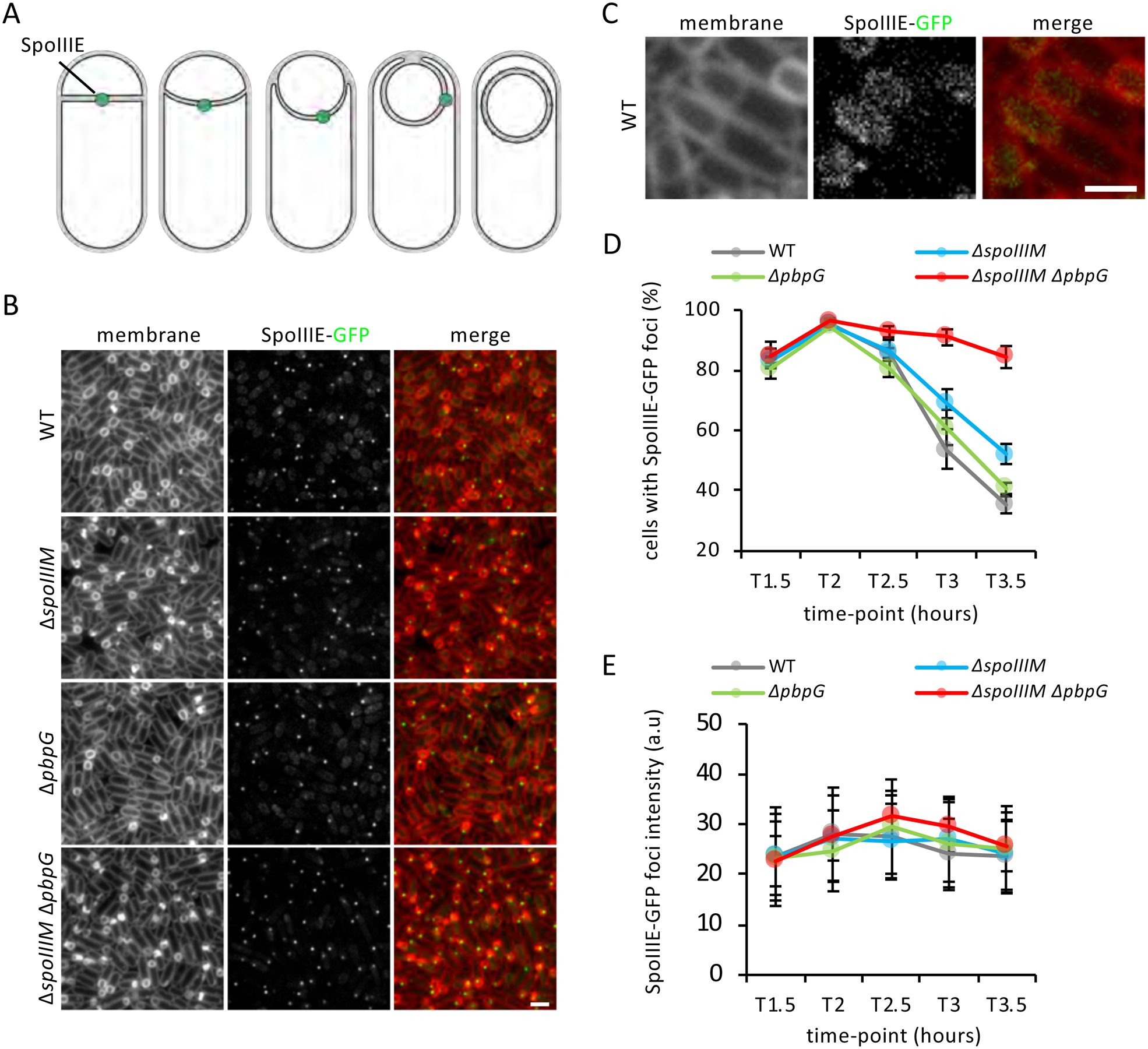
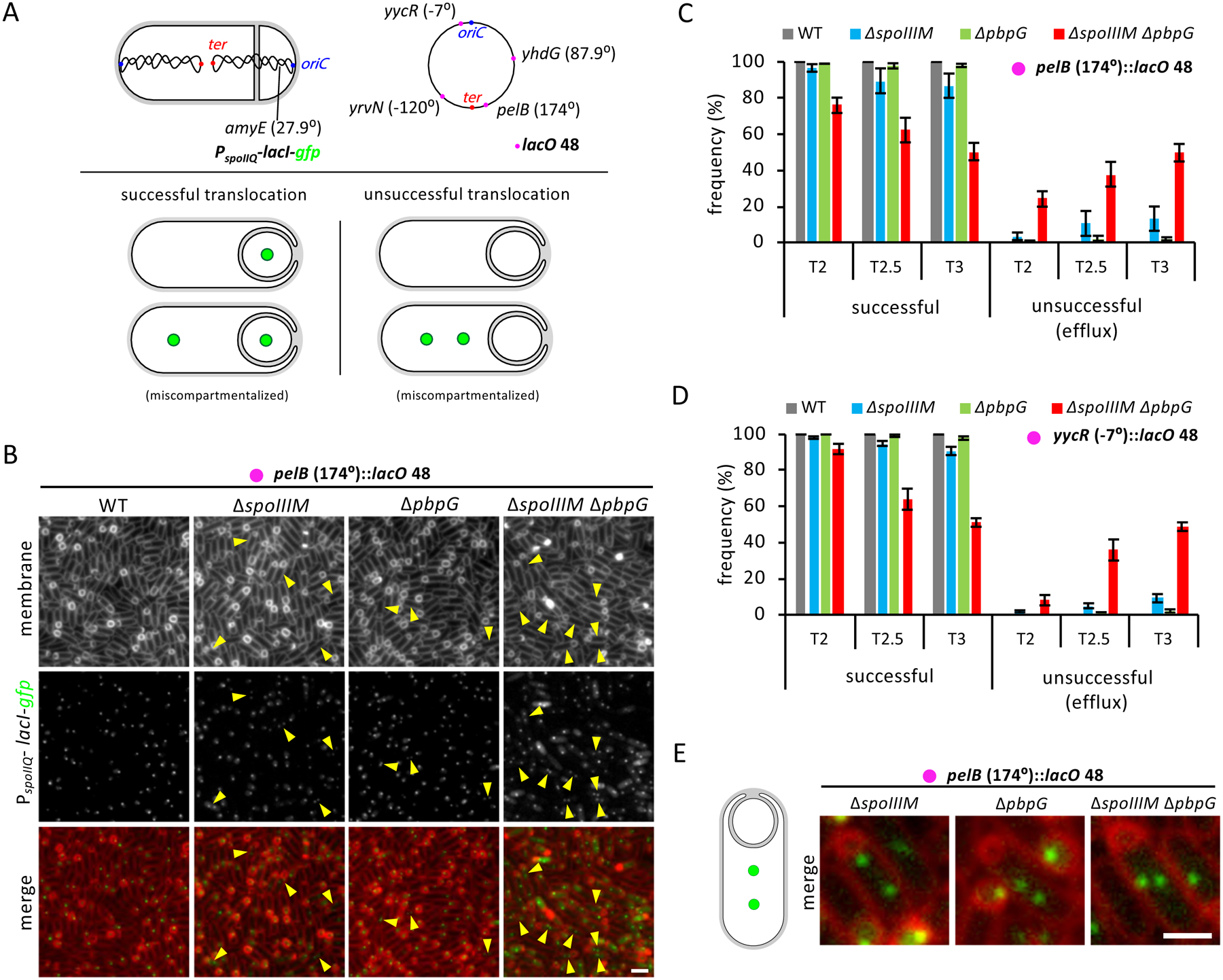
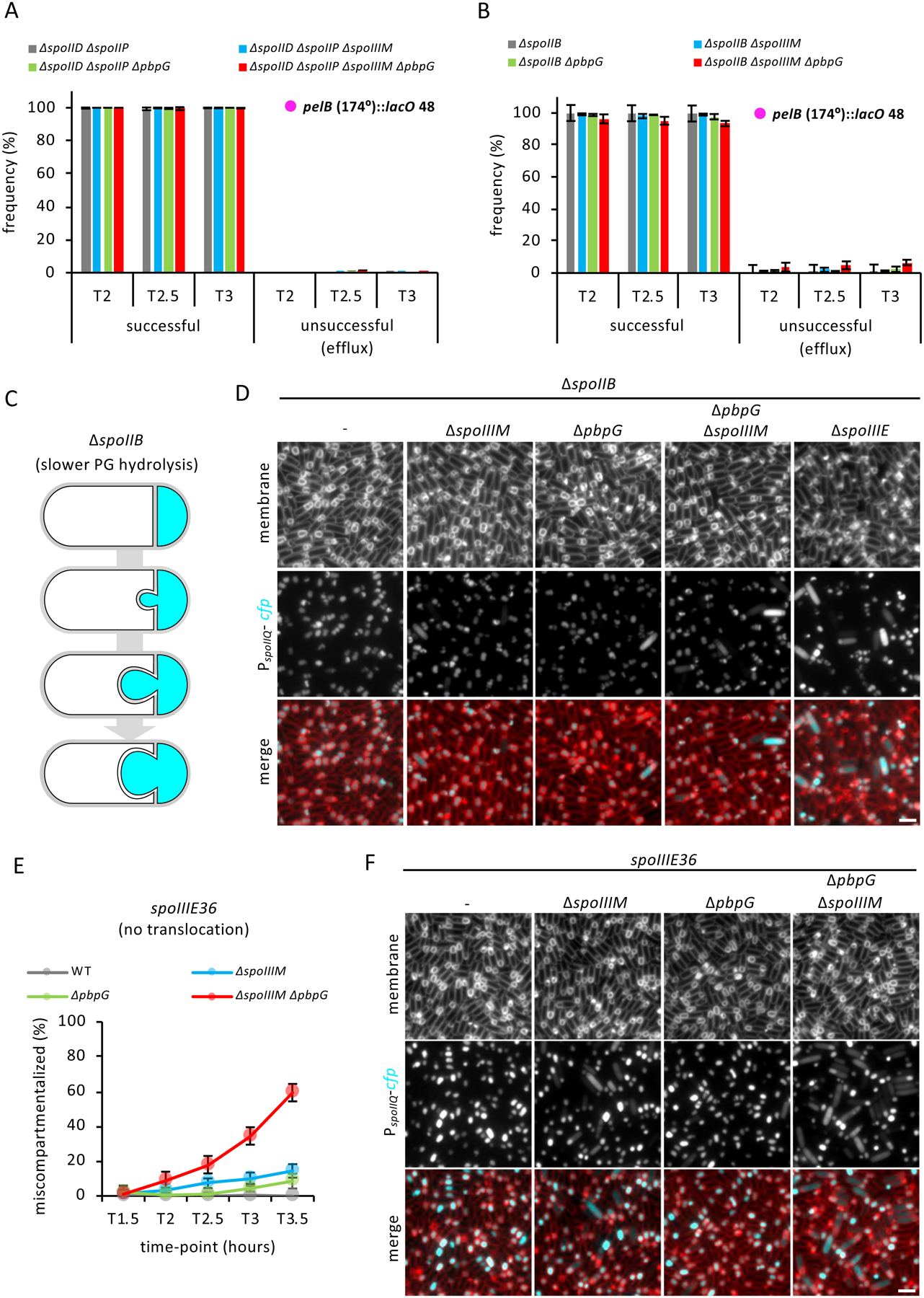
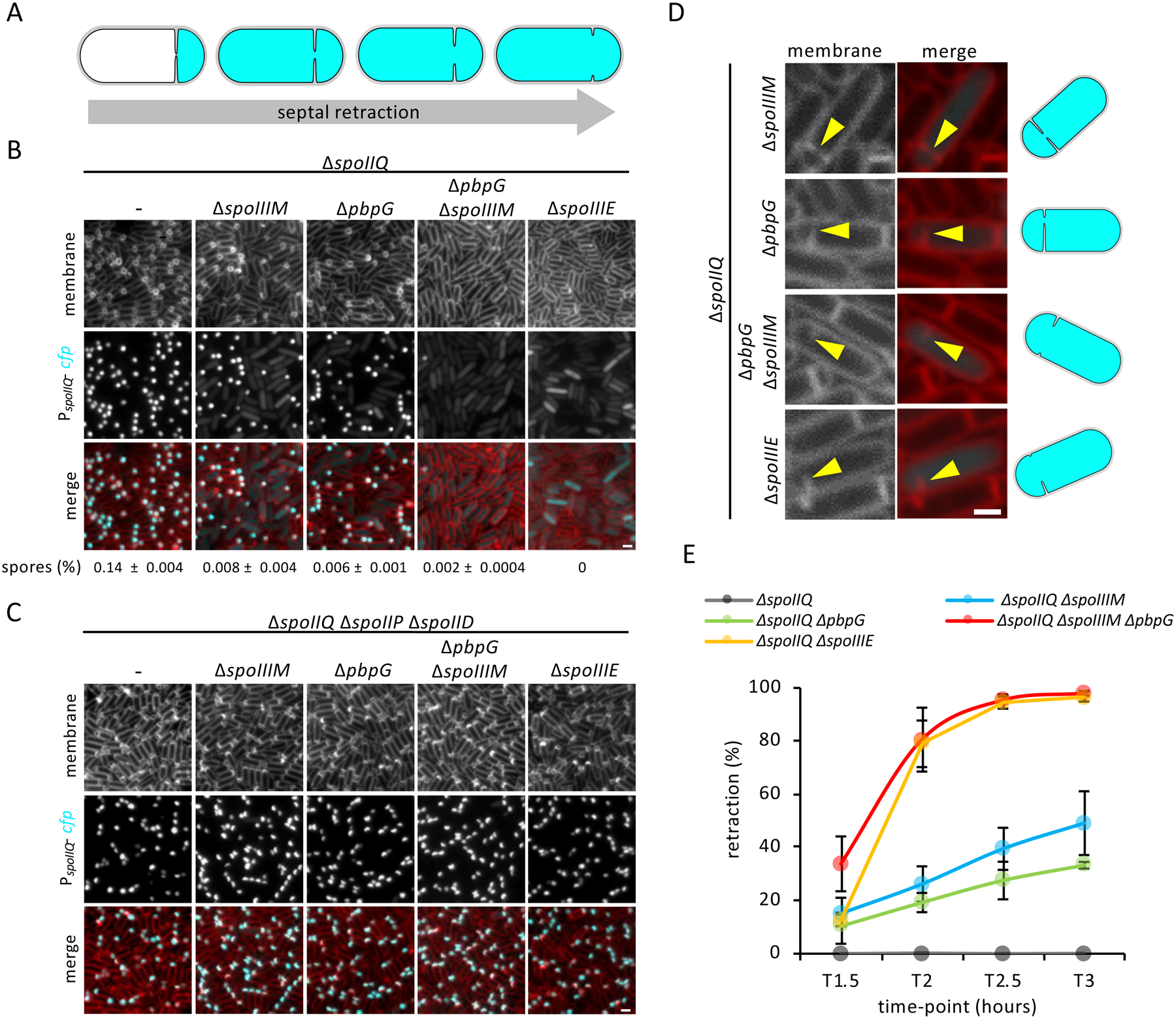
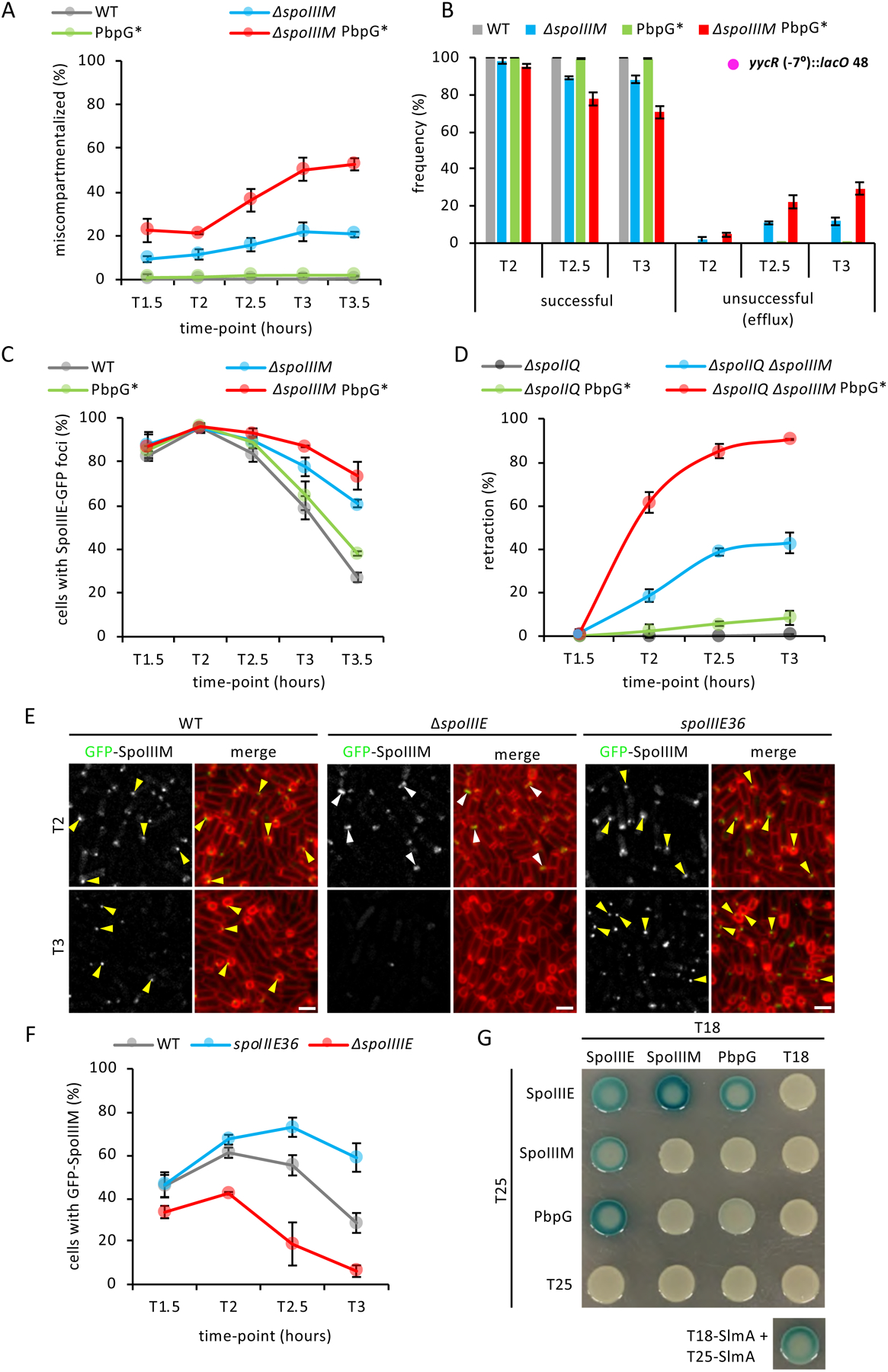
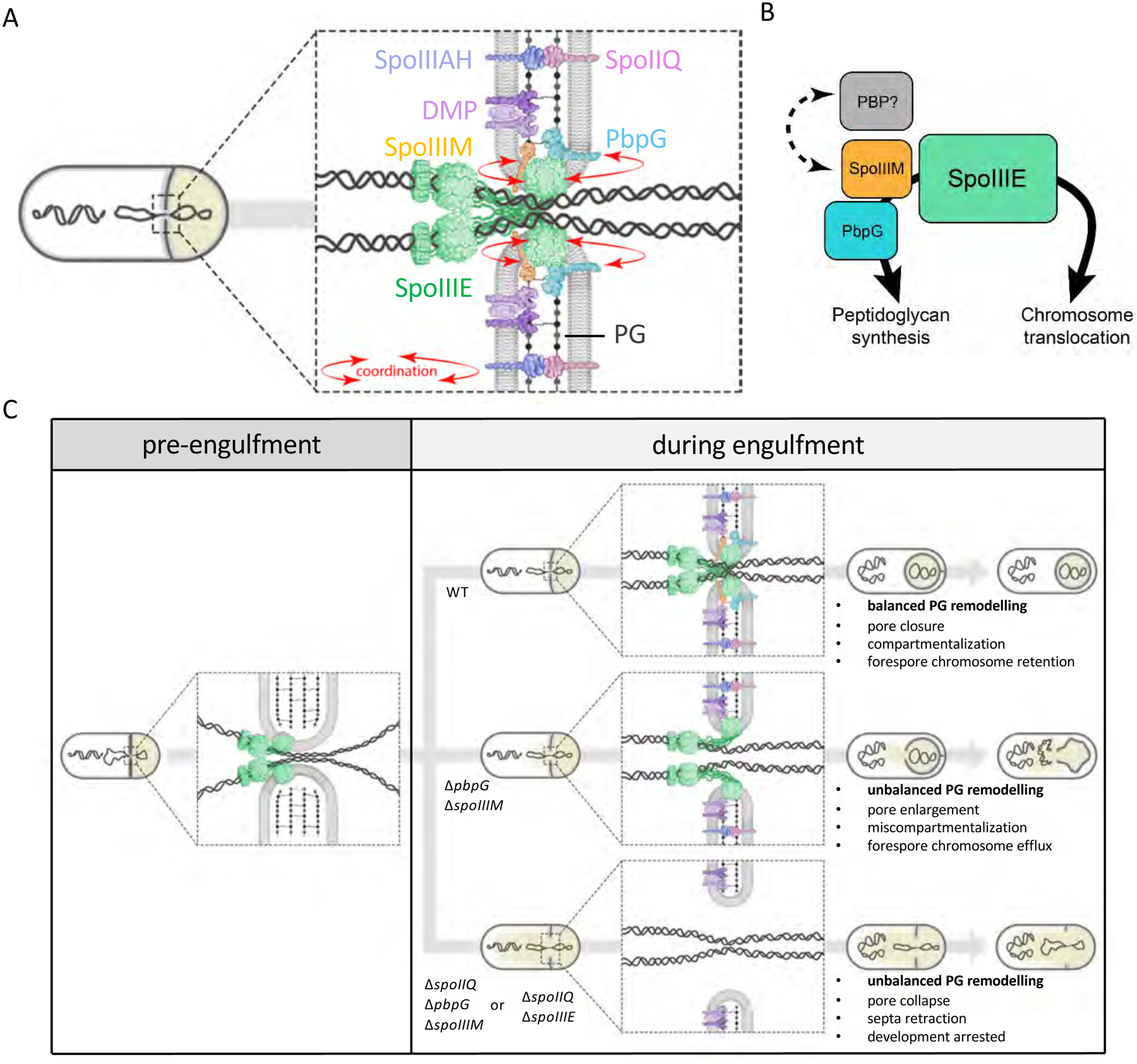
Asymmetric division, a hallmark of endospore development, generates two cells, a larger mother cell and a smaller forespore. Approximately 75% of the forespore chromosome must be translocated across the division septum into the forespore by the DNA translocase SpoIIIE. Asymmetric division also triggers cell-specific transcription, which initiates septal peptidoglycan remodeling involving synthetic and hydrolytic enzymes. How these processes are coordinated has remained a mystery. Using Bacillus subtilis, we identified factors that revealed the link between chromosome translocation and peptidoglycan remodeling. In cells lacking these factors, the asymmetric septum retracts, resulting in forespore cytoplasmic leakage and loss of DNA translocation. Importantly, these phenotypes depend on septal peptidoglycan hydrolysis. Our data support a model in which SpoIIIE is anchored at the edge of a septal pore, stabilized by newly synthesized peptidoglycan and protein-protein interactions across the septum. Together, these factors ensure coordination between chromosome translocation and septal peptidoglycan remodeling to maintain spore development.
Keywords: SpoIIIE; cell wall; chromosome segregation; chromosome translocation; development; endospores; peptidoglycan; spores; sporulation.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
