

Cross-seeding between the functional amyloidogenic CRES and CRES3 family members and their regulation of Aβ assembly
- PMID: 33384380
- PMCID: PMC7948811
- DOI: 10.1074/jbc.RA120.015307
Cross-seeding between the functional amyloidogenic CRES and CRES3 family members and their regulation of Aβ assembly
Abstract
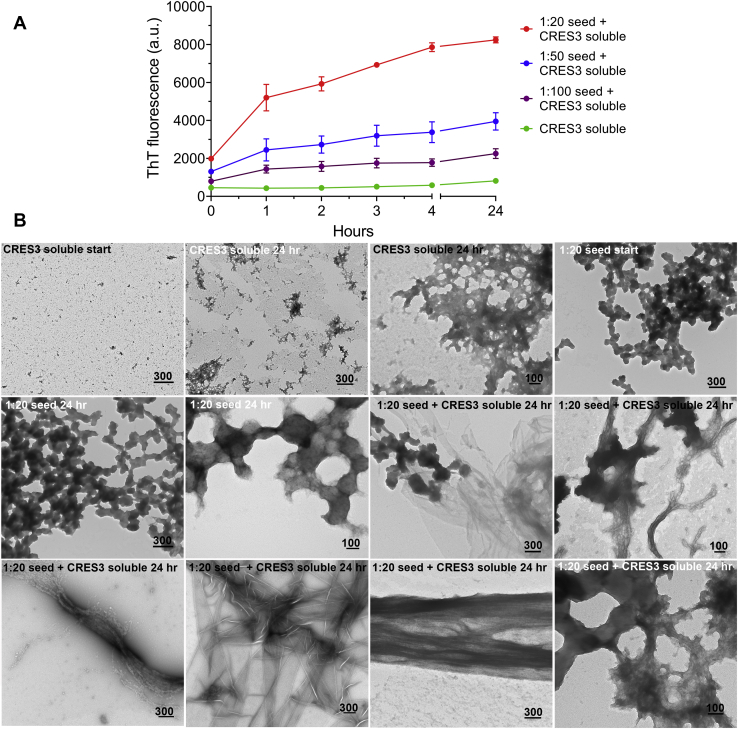
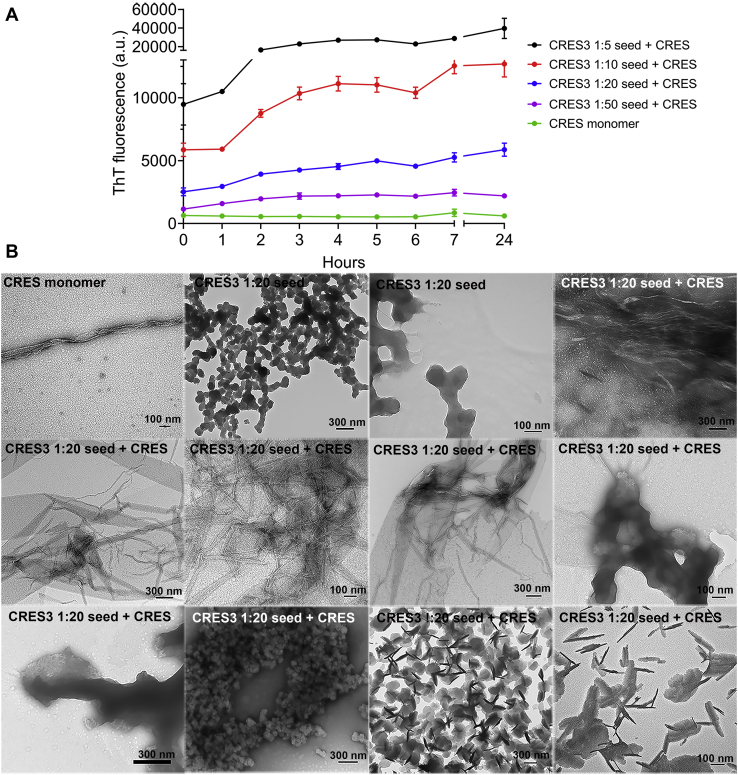
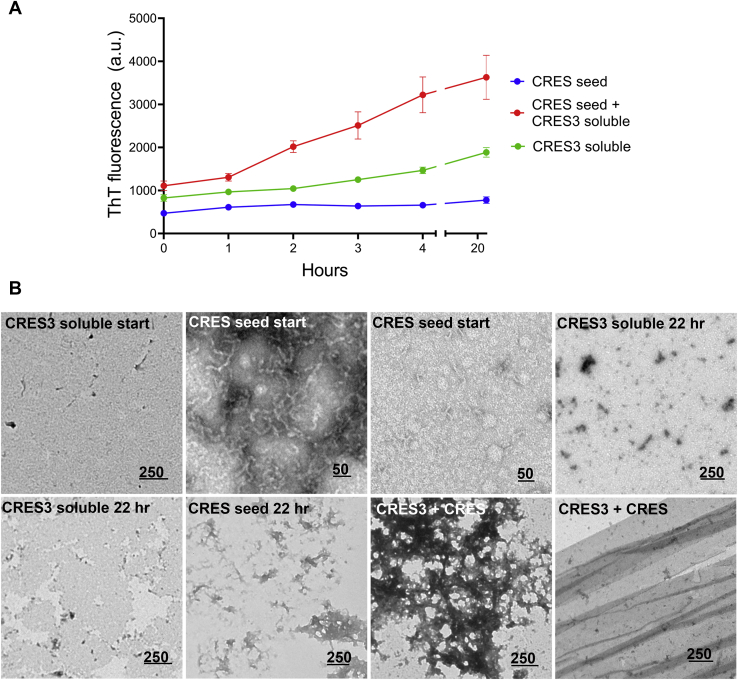
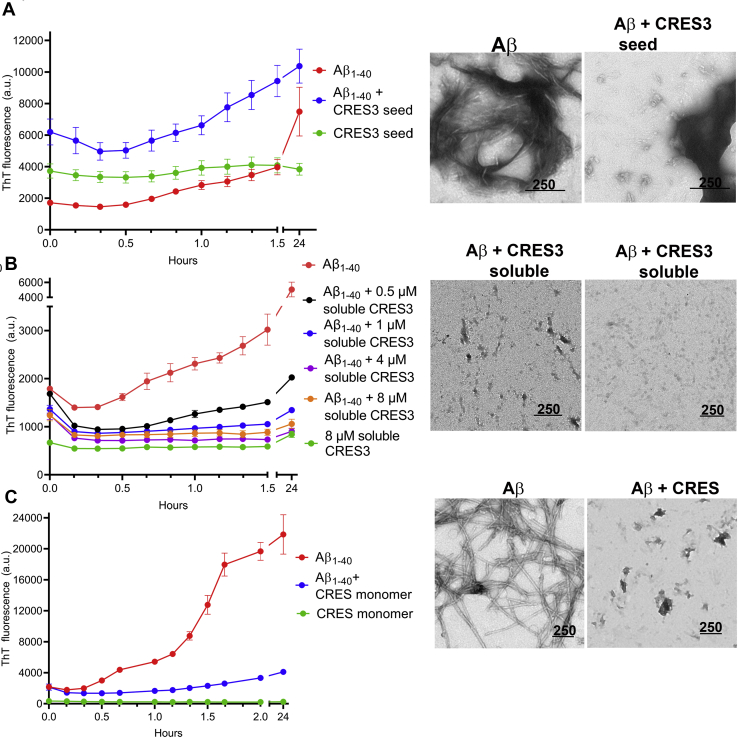
Accumulating evidence shows that amyloids perform biological roles. We previously showed that an amyloid matrix composed of four members of the CRES subgroup of reproductive family 2 cystatins is a normal component of the mouse epididymal lumen. The cellular mechanisms that control the assembly of these and other functional amyloid structures, however, remain unclear. We speculated that cross-seeding between CRES members could be a mechanism to control the assembly of the endogenous functional amyloid. Herein we used thioflavin T assays and negative stain transmission electron microscopy to explore this possibility. We show that CRES3 rapidly formed large networks of beaded chains that possessed the characteristic cross-β reflections of amyloid when examined by X-ray diffraction. The beaded amyloids accelerated the amyloidogenesis of CRES, a less amyloidogenic family member, in seeding assays during which beads transitioned into films and fibrils. Similarly, CRES seeds expedited CRES3 amyloidogenesis, although less efficiently than the CRES3 seeding of CRES. These studies suggest that CRES and CRES3 hetero-oligomerize and that CRES3 beaded amyloids may function as stable preassembled seeds. The CRES3 beaded amyloids also facilitated assembly of the unrelated amyloidogenic precursor Aβ by providing a surface for polymerization though, intriguingly, CRES3 (and CRES) monomer/early oligomer profoundly inhibited Aβ assembly. The cross-seeding between the CRES subgroup members is similar to that which occurs between bacterial curli proteins suggesting that it may be an evolutionarily conserved mechanism to control the assembly of some functional amyloids. Further, interactions between unrelated amyloidogenic precursors may also be a means to regulate functional amyloid assembly.
Keywords: Aβ; CRES subgroup; amyloid; cross-seeding; cystatin; epididymis; mouse.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
